多组学解析KDM5C、KDM6A与KMT2B在癌症表观遗传失调与转录重编程中的作用机制
《Communications Biology》:Multi-omics elucidation of KDM5C, KDM6A, and KMT2B roles in cancer epigenetic dysregulation and transcriptional reprogramming
【字体:
大
中
小
】
时间:2025年12月04日
来源:Communications Biology 5.1
编辑推荐:
本研究通过整合多组学技术,系统阐明了组蛋白修饰酶KDM5C、KDM6A和KMT2B在癌症表观遗传调控中的关键作用。研究人员利用CRISPR/Cas9基因编辑技术构建敲除细胞模型,结合ATAC-seq、染色质结合蛋白质组学和转录组学分析,发现这三种酶通过调控染色质可及性和转录因子结合,影响癌相关信号通路。特别验证了KDM6A缺失通过改变CDH家族基因表达促进胰腺癌细胞迁移侵袭。该研究为理解表观遗传失调在肿瘤发生中的机制提供了新视角,为开发靶向治疗策略奠定了理论基础。
在肿瘤生物学研究领域,表观遗传调控机制日益受到关注。组蛋白修饰作为表观遗传调控的重要方式,通过动态改变染色质结构影响基因表达模式。组蛋白甲基化修饰尤其关键,例如H3K4me3(组蛋白H3第4位赖氨酸三甲基化)通常标记活跃启动子区域,而H3K27me3(组蛋白H3第27位赖氨酸三甲基化)则与基因沉默相关。这些修饰的平衡由组蛋白修饰酶(HME)精密调控,包括甲基转移酶和去甲基化酶。然而,在癌症中,组蛋白修饰酶经常发生突变,导致表观遗传景观紊乱,进而驱动肿瘤发生发展。
一个令人困惑的现象是,功能相反的组蛋白修饰酶在多种癌症中同时存在突变。例如,KMT2B(H3K4me3甲基转移酶)和KDM5C(H3K4me3去甲基化酶)在超过10%的鳞状细胞癌中发生突变,而KDM6A(H3K27me3去甲基化酶)在超过25%的尿路上皮癌中发生改变。这些共存突变提示肿瘤发生机制可能超越了经典的组蛋白标记稳态调控。事实上,组蛋白修饰酶突变可能产生谱系特异性甚至矛盾的效果,这表明它们可能在更广泛的转录因子驱动网络中发挥作用,但这些网络的具体组成和层级关系仍不明确。
为了解决这一科学问题,上海交通大学的研究团队在《Communications Biology》上发表了最新研究成果。他们采用多组学整合分析框架,在统一的细胞背景下系统研究了KDM5C、KDM6A和KMT2B这三种组蛋白修饰酶的功能。
研究人员主要运用了CRISPR/Cas9基因编辑技术构建敲除细胞模型,通过ATAC-seq(转座酶可及染色质测序)分析全基因组染色质可及性变化,利用染色质结合蛋白质组学鉴定蛋白质-DNA相互作用,并采用RNA-seq(转录组测序)评估基因表达谱改变。此外,还通过蛋白质印迹、细胞功能实验和生物信息学分析等多种技术手段验证研究结果。
KDM5C、KDM6A和KMT2B在不同肿瘤中显示高突变频率
研究人员首先通过生物信息学分析证实了KDM5C、KDM6A和KMT2B在多种人类恶性肿瘤中具有高频遗传变异。这些酶在特定癌症类型中位列前20个最常突变基因,其中大多数突变为截短或错义变异,提示功能失活。表达模式分析显示,去甲基化酶KDM5C和KDM6A在多数癌症类型中表达下调,而甲基转移酶KMT2B则在多种恶性肿瘤中表达上调。临床预后分析表明,KDM5C和KDM6A的低表达与膀胱尿路上皮癌患者不良生存结局相关,而KMT2B的高表达则预示肾上腺皮质癌患者预后较差。
CRISPR介导的KDM5C、KDM6A和KMT2B敲除改变组蛋白修饰并驱动细胞表型重编程
研究团队利用CRISPR/Cas9基因组编辑技术成功构建了KDM5C、KDM6A和KMT2B敲除的HEK293T细胞系。蛋白质印迹分析验证了这些酶对组蛋白修饰的特异性影响:KDM5C敲除细胞中H3K4me3水平升高,KMT2B敲除细胞中H3K4me3水平降低,而KDM6A敲除细胞中H3K27me3水平增加。细胞功能实验显示,三种酶的敲除均一致抑制细胞增殖,但只有KDM6A缺失增强细胞迁移能力,KDM5C或KMT2B缺失对细胞迁移无明显影响。
CRISPR介导的KDM5C、KDM6A和KMT2B敲除诱导全基因组染色质可及性改变
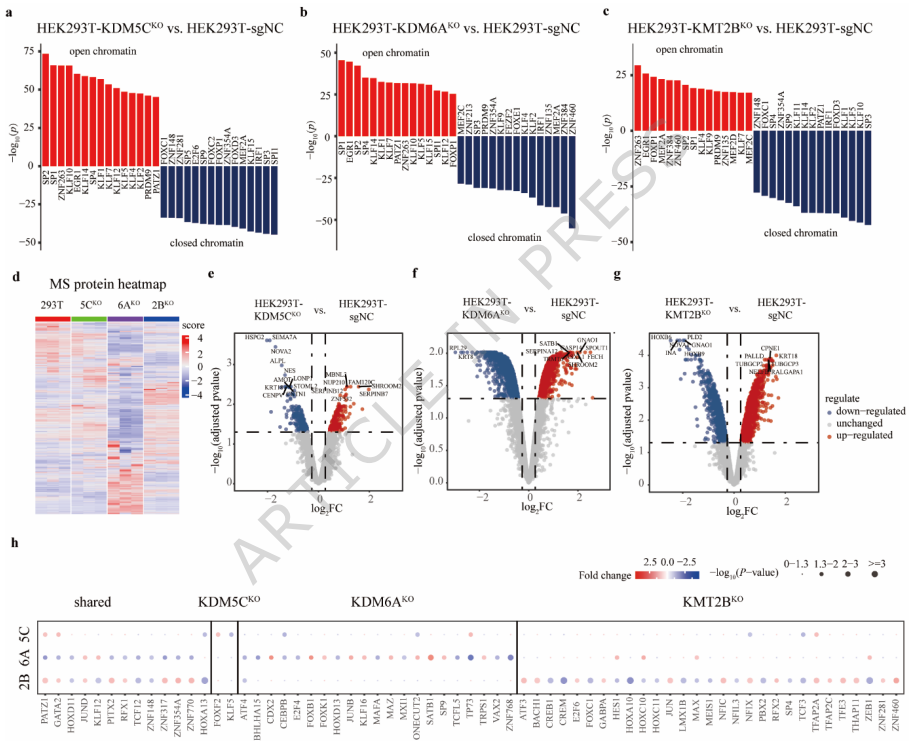
ATAC-seq分析揭示了三种敲除细胞系中全基因组染色质可及性的显著变化。KDM5C敲除导致约2,029个位点发生可及性改变,其中70%为可及性增加;KDM6A缺失影响2,077个位点,73%为可及性降低;KMT2B敲除引起2,753个位点变化,58%为可及性降低。差异可及区域主要位于启动子、内含子、基因间区和外显子区域。值得注意的是,KDM5C缺失增强启动子区域可及性,与H3K4me3水平升高一致;KMT2B缺失降低启动子可及性,反映H3K4me3水平下降;KDM6A缺失则降低启动子和内含子区域可及性,与H3K27me3水平增加相符。
KDM5C、KDM6A和KMT2B敲除细胞中的染色质蛋白质组重塑揭示染色质动力学和转录调控改变
染色质结合蛋白质组学分析显示,三种敲除细胞系均发生显著的蛋白质组重塑。KDM5C敲除细胞中鉴定出4,610个染色质结合蛋白,其中248个下调,226个上调;KDM6A敲除细胞中检测到4,820个蛋白,836个下调,733个上调;KMT2B敲除细胞中发现4,560个蛋白,848个下调,1,022个上调。KEGG(京都基因与基因组百科全书)通路富集分析表明,这些差异蛋白主要富集于细胞周期调控、激素信号和感染相关通路,提示三种酶的缺失主要干扰转录调控过程。
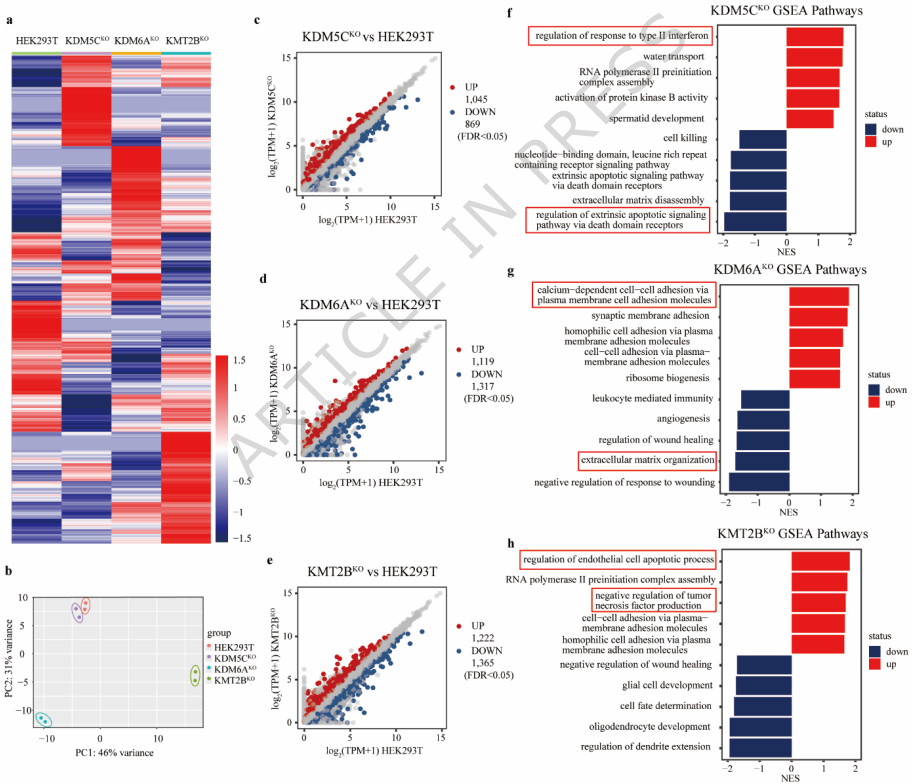
KDM5C、KDM6A和KMT2B缺陷导致的转录组失调
转录组分析发现,KDM5C敲除导致1,045个基因上调和869个基因下调;KDM6A敲除引起1,119个基因上调和1,317个基因下调;KMT2B敲除导致1,222个基因上调和1,365个基因下调。基因集富集分析(GSEA)显示,敲除细胞中多个癌症相关信号通路显著富集,包括II型干扰素反应调控、细胞间粘附和内皮细胞凋亡过程调控。此外,KMT2B敲除引起多个相关组蛋白修饰酶(如KDM5A/B/C、KDM6A和KMT2C/D)的代偿性上调。
KDM5C、KDM6A和KMT2B敲除细胞中通过染色质结合蛋白质组失调进行的致癌转录重编程
整合分析显示,染色质可及性改变与基因表达模式之间存在明显相关性:上调基因与染色质开放增加相关,而下调基因与染色质关闭相关。研究人员通过比较启动子区域差异可及性峰预测的转录因子和染色质结合蛋白质组鉴定的差异表达蛋白,发现每种敲除细胞都具有特定的转录因子调控网络。KDM5C敲除细胞中,FOXF2和KLF5等转录因子发生特异性改变;KDM6A敲除影响KLF16、TP73和JUNB等因子;KMT2B敲除则调控JUN、NFIC和HOXA10等因子。值得注意的是,PATZ1和GATA2等转录因子在三种敲除细胞中均发生改变。
KDM6A缺失改变细胞粘附基因表达,增强PANC-1细胞迁移和侵袭
鉴于KDM6A敲除细胞显示细胞迁移增强和细胞间粘附通路富集,研究人员检测了多个细胞粘附基因的mRNA水平。发现CDH2和CDH12表达上调,而CDH3表达下调。相应地,CDH2和CDH12的启动子区域染色质可及性增加,CDH3启动子区域可及性降低。在胰腺癌PANC-1细胞中敲低KDM6A后,细胞迁移和侵袭能力增强,CDH2、CDH3和CDH12表达水平也发生类似改变。
本研究通过多组学整合分析,系统阐明了KDM5C、KDM6A和KMT2B在癌症表观遗传调控中的关键作用。研究发现这三种组蛋白修饰酶通过独特而相互关联的机制调控染色质可及性、转录因子结合动力学和致癌基因表达程序。与预期相反,KMT2B缺失(H3K4me3耗竭)和KDM5C缺失(H3K4me3积累)并未诱导相反的转录程序,KDM6A缺陷(H3K27me3积累)也未表现出独特的调控效应。相反,每种酶的调控都产生独特的影响,体现在转录输出和染色质相关蛋白质组状态两个方面。
功能上,KDM5C缺失上调FOXF2并下调KLF5,涉及G蛋白偶联受体通路失调;KDM6A缺失上调JUNB并下调TP73,影响细胞外基质调控;KMT2B缺失上调JUN并下调HOXA10,影响细胞因子信号传导。值得注意的是,PATZ1和GATA2等转录因子在三种敲除细胞中均发生改变。在PANC-1胰腺癌细胞中,进一步证实KDM6A通过调控控制细胞粘附的CDH家族基因,从而促进迁移和侵袭。
整合分析证明了启动子可及性、转录因子占据和基因表达之间的强相关性,并揭示了表观遗传和遗传驱动因子之间的协同作用。这些发现揭示了组蛋白修饰酶在上下文依赖的功能层级,强调了多层次分析对于解析癌症表观遗传调控复杂性的必要性。该研究为理解表观遗传失调在肿瘤发生中的机制提供了新视角,为开发靶向治疗策略奠定了理论基础。
 生物通微信公众号
生物通微信公众号
 生物通新浪微博
生物通新浪微博
今日动态 |
人才市场 |
新技术专栏 |
中国科学人 |
云展台 |
BioHot |
云讲堂直播 |
会展中心 |
特价专栏 |
技术快讯 |
免费试用
版权所有 生物通
Copyright© eBiotrade.com, All Rights Reserved
联系信箱:
粤ICP备09063491号