恶性疟原虫DNA修复动态揭示TLS聚合酶与PfRad51在基因组多样化中的独特作用
《Nucleic Acids Research》:Plasmodium falciparum DNA repair dynamics reveal unique roles for TLS polymerases and PfRad51 in genome diversification
【字体:
大
中
小
】
时间:2025年11月25日
来源:Nucleic Acids Research 13.1
编辑推荐:
本刊推荐:为解决恶性疟原虫基因组核心区保守而亚端粒区高度变异的分子机制问题,研究人员聚焦DNA修复通路动态差异,发现PfRad51缺失导致核心区同源重组(HR)修复完全失效,而亚端粒区存在独立于PfRad51和TLS聚合酶(PfPolζ/PfRev1)的新型HR通路。该研究首次揭示寄生虫通过区域特异性修复策略维持基因组二象性,为理解抗原变异机制提供新视角。
在人类与疟疾的漫长斗争中,恶性疟原虫(Plasmodium falciparum)始终以其惊人的变异能力挑战着全球公共卫生系统。这种单细胞寄生虫拥有一个奇特的基因组结构:14条染色体被清晰地划分为高度保守的核心区域和充满变数的亚端粒区域。后者聚集着多拷贝基因家族(如var、rifin、stevor等),其中var基因编码的PfEMP1蛋白更是寄生虫躲避宿主免疫攻击的关键武器。然而,这种基因组"二元性"的维持机制一直是未解之谜――为何寄生虫能在亚端粒区疯狂"洗牌"基因的同时,却让核心基因组保持稳定?
更令人困惑的是,恶性疟原虫的生物学特性似乎与常规DNA修复逻辑相悖:作为单倍体生物,它缺乏经典的非同源末端连接(C-NHEJ)修复通路,却要依赖同源重组(HR)来修复致命的DNA双链断裂(DSBs)。在缺乏同源染色体模板的情况下,它如何完成高保真修复?近年来研究提示转损合成(TLS)聚合酶和HR关键蛋白PfRad51可能参与变异过程,但二者在时空特异性修复中的作用仍是空白。正是这些矛盾与谜题,促使Weill Cornell医学中心的研究团队在《Nucleic Acids Research》上发表了这项揭示疟原虫DNA修复动态的研究。
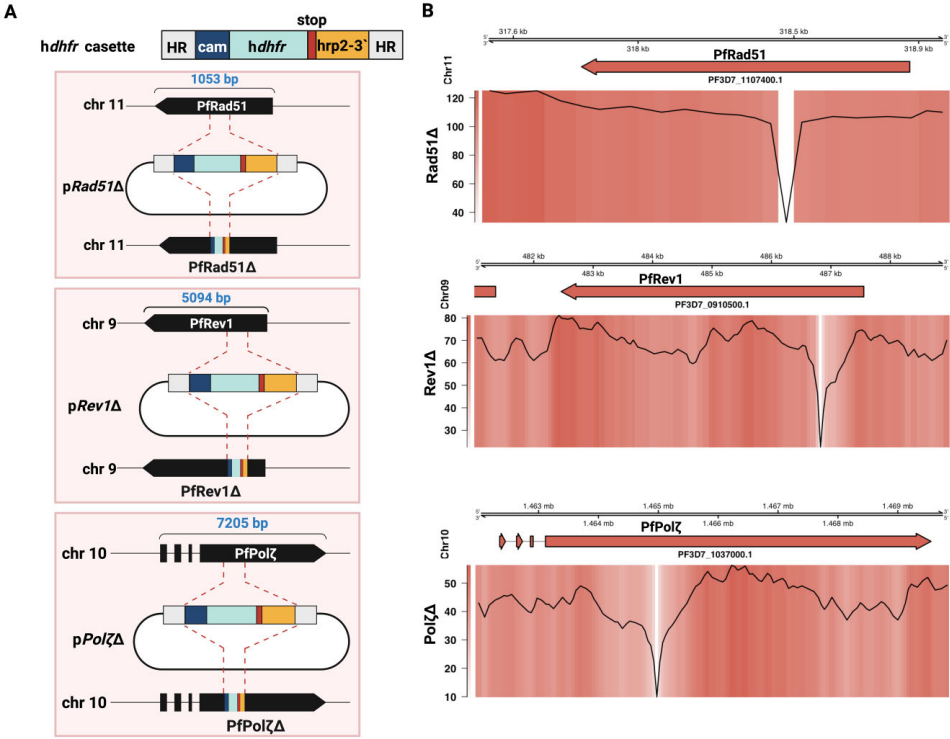
研究人员首先通过CRISPR/Cas9基因编辑技术成功构建了PfRad51、PfRev1和PfPolζ的基因敲除株系(图1)。令人意外的是,这三种在高等生物中往往至关重要的修复蛋白的缺失,并未影响寄生虫在正常培养条件下的增殖能力(补充图S2C)。
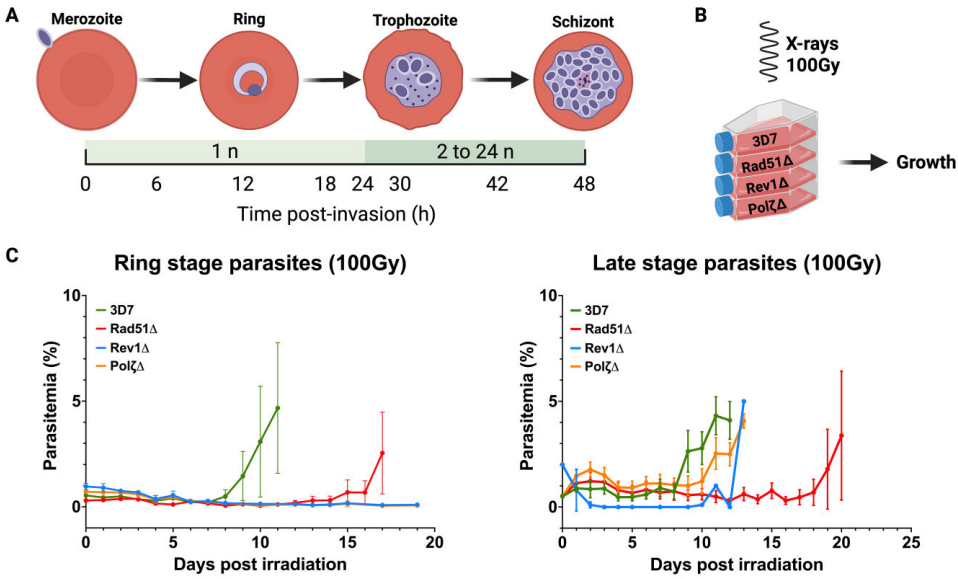
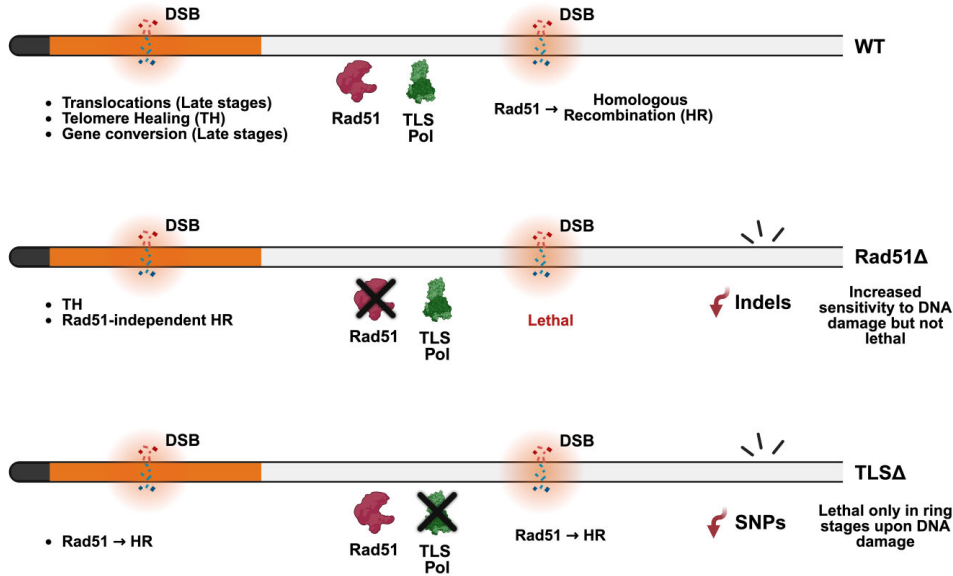
当研究团队用X射线(100 Gy)诱导全局DNA损伤时,戏剧性的阶段特异性差异浮出水面(图2)。在复制活跃的晚期滋养体阶段,所有突变株都能在15-19天内恢复生长,表明HR通路的主导作用。然而在单倍体的环状体阶段,TLS聚合酶敲除株完全丧失修复能力,而PfRad51敲除株仍能存活(图2C)。这一发现首次证明TLS通路是环状体阶段修复的"生命线",而PfRad51非HR通路在晚期阶段具有补偿潜力。
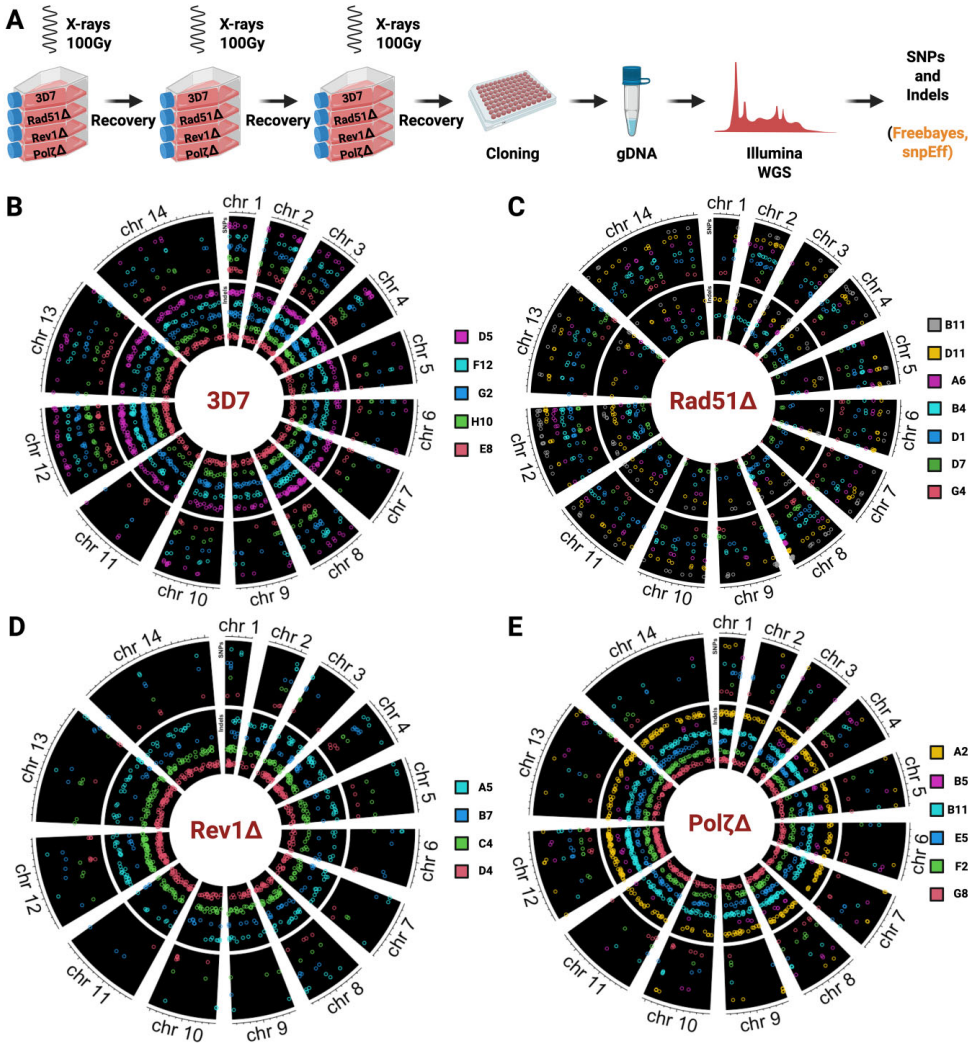
全基因组测序进一步揭示了突变谱的差异(图3)。TLS敲除株表现出全面的"低突变"表型,SNP数量显著减少(图4A),符合其经典易错修复特性。而PfRad51敲除株则出现矛盾现象:编码区非同义SNP增加(图4C),但插入缺失(indel)频率大幅降低(图4D)。更值得注意的是,野生型寄生虫的突变优先富集在亚端粒区(图4B),这种区域偏好性在TLS突变株中减弱,暗示TLS聚合酶是亚端粒变异的重要推手。
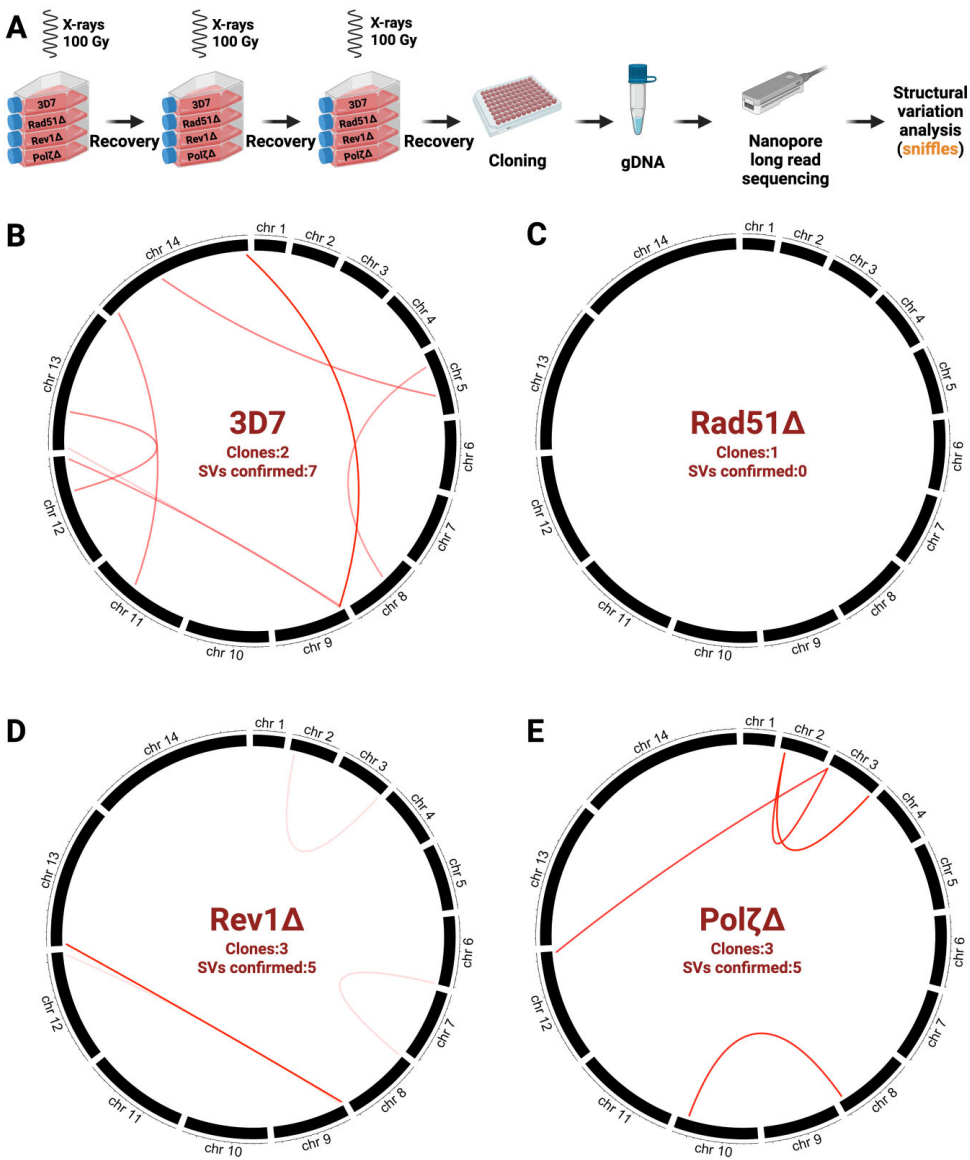
纳米孔测序捕捉到的染色体易位事件更凸显了PfRad51的核心地位(图5)。野生型与TLS突变株平均每个克隆产生3-4次亚端粒区易位,而PfRad51敲除株则完全未检测到易位事件(图5B-C)。这表明尽管PfRad51非依赖通路能保证基础存活,但规范的HR仍是基因组重排的主要驱动力。
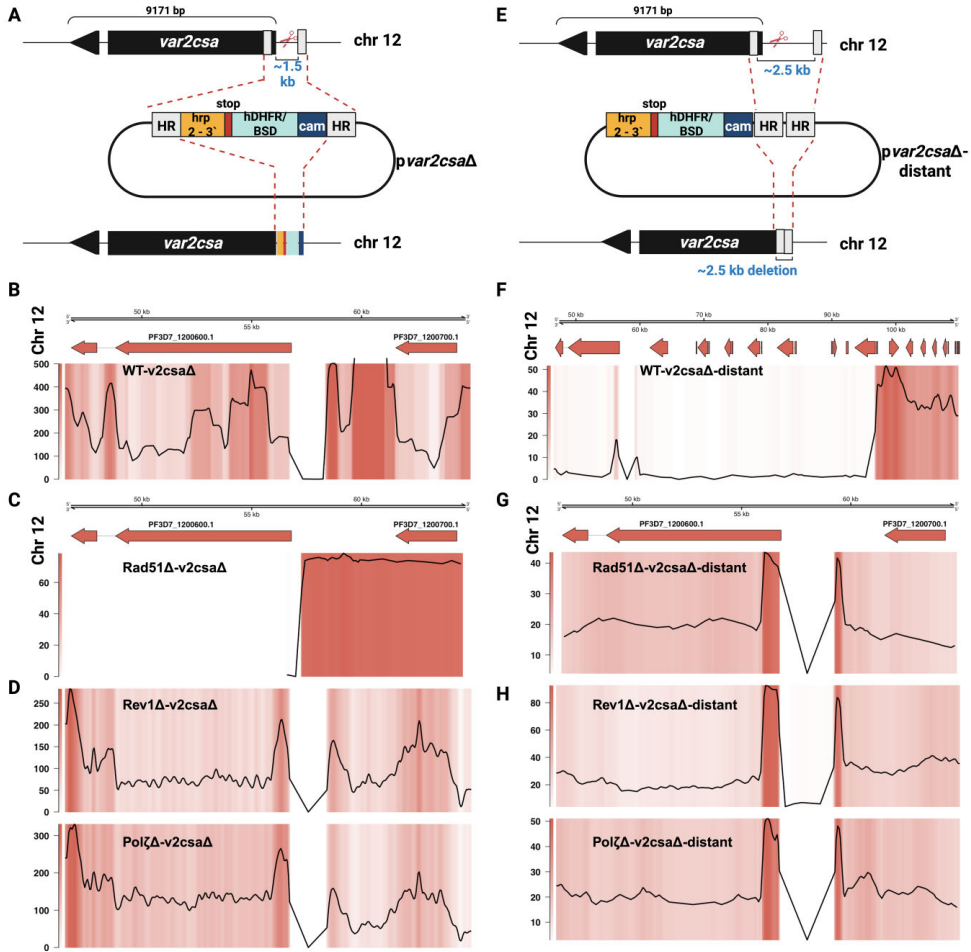
最精彩的发现来自针对特定基因座的CRISPR/Cas9修复实验(图6)。当在亚端粒区var2csa基因附近引入DSB时,野生型寄生虫能有效利用同源模板进行HR修复(图6B)。而PfRad51敲除株原本只能通过末端愈合(TH)机制修复――直到研究人员将同源臂间距增至2.5kb后,竟意外激活了全新的PfRad51非依赖型HR通路(图6G)。这种新型修复表现出异常模式:修复DNA多次复制质粒骨架后才完成重组(补充图S6),且在所有三种突变株中均能发生(图6H)。
与之形成鲜明对比的是核心基因组的结果(图7)。当靶向非必需基因PfUPF1时,PfRad51的缺失直接导致DSB修复失败,证明经典HR通路在核心区不可替代。而TLS聚合酶在此过程中完全冗余(图7C,G)。这种"地域歧视"般的修复策略,完美解释了寄生虫如何实现基因组二元性控制。
这项研究通过整合基因编辑(CRISPR/Cas9)、阶段特异性辐照实验、双平台全基因组测序(Illumina短读长与Oxford Nanopore长读长)以及定位修复分析等技术手段,系统描绘了恶性疟原虫DNA修复网络的时空动态图谱。研究不仅证实了TLS聚合酶在单倍体阶段的不可替代性,更揭示了PfRad51在indel形成和染色体易位中的主导作用。而最突破性的发现在于亚端粒区存在的PfRad51非依赖型HR通路,这为解释var基因多样性爆发提供了全新机制(图8)。
该研究颠覆了我们对单倍体生物DNA修复能力的认知,证明恶性疟原虫进化出了远比预期复杂的修复策略。这种区域特异性的修复"双轨制"既确保了核心基因组的稳定,又为亚端粒抗原基因的持续进化开辟了通道。未来对新型HR通路元件的鉴定,不仅可能揭示疟原虫免疫逃逸的终极密码,更为开发阻断抗原变异的新型抗疟策略提供了独特靶点。在疟原虫与人类宿主这场持续千年的"军备竞赛"中,这项研究为我们打开了窥见敌方武器库的新窗口。
 生物通微信公众号
生物通微信公众号
 生物通新浪微博
生物通新浪微博
今日动态 |
人才市场 |
新技术专栏 |
中国科学人 |
云展台 |
BioHot |
云讲堂直播 |
会展中心 |
特价专栏 |
技术快讯 |
免费试用
版权所有 生物通
Copyright© eBiotrade.com, All Rights Reserved
联系信箱:
粤ICP备09063491号