DNA双链断裂簇通过抑制经典非同源末端连接和促进替代性末端连接显著增强基因突变频率
《Scientific Reports》:CRISPR/Cas9 generated DSB clusters mimic complex lesions induced by high-LET radiation and shift repair from c-NHEJ to mutagenic repair pathways
【字体:
大
中
小
】
时间:2025年10月21日
来源:Scientific Reports 3.9
编辑推荐:
本研究针对高线性能量转移辐射诱导的复杂DNA双链断裂(DSB)簇难以模拟的问题,利用CRISPR/Cas9系统在hHPRT基因位点精确构建单DSB与DSB簇模型。研究发现DSB簇通过抑制DNA-PKcs依赖的经典非同源末端连接(c-NHEJ),激活PARP1和LIG3介导的替代性末端连接(alt-EJ),显著提升突变频率,揭示了DSB复杂度作为决定修复路径选择与基因组不稳定性的关键因素。该成果为高LET辐射的遗传风险评估提供了新视角。
当我们仰望星空,或许不会想到那些穿越宇宙的高能粒子同样在微观世界掀起波澜。高线性能量转移(高LET)辐射,如重离子或α粒子,因其在肿瘤放疗中的深度穿透优势和相对生物学效应(RBE)而备受关注。然而,这类辐射在杀死癌细胞的同时,也会在DNA上留下独特的"伤痕"――复杂DNA双链断裂(DSB)簇,即多个DSB紧密聚集在狭小基因组区域内。与传统辐射产生的"干净"DSB不同,这些DSB簇如同DNA上的"车祸现场",更难被准确修复,更易导致基因突变、染色体畸变甚至细胞癌变。
尽管其重要性不言而喻,如何在实验室中精确模拟这些DSB簇并揭示其修复机制,一直是辐射生物学领域的难题。现有的随机照射方法无法控制DSB的数量、位置和复杂度,而定制化DSB簇的生成技术则鲜有报道。正是为了攻克这一瓶颈,由Emil Mladenov领衔的研究团队在《Scientific Reports》上发表了创新性研究,他们巧妙利用CRISPR/Cas9基因编辑技术,在人类hHPRT基因位点上"精雕细琢",首次系统比较了单DSB与不同复杂度DSB簇的突变诱导能力,并深入揭示了其背后的分子通路"博弈"。
研究团队主要运用了以下关键技术:首先,通过CRISPR/Cas9系统在A549细胞系hHPRT基因位点精确引入单DSB或DSB簇;其次,采用6-硫代鸟嘌呤(6TG)抗性筛选和集落形成实验量化突变频率(MF);再者,利用蛋白质印迹(Western blot)和间接免疫荧光技术验证关键蛋白表达与定位;此外,通过Surveyor核酸酶检测和T7E1酶切分析验证DSB形成效率;最后,结合药理学抑制和基因敲除(如DNA-PKcs、PARP1、ATM)模型解析特定通路功能。
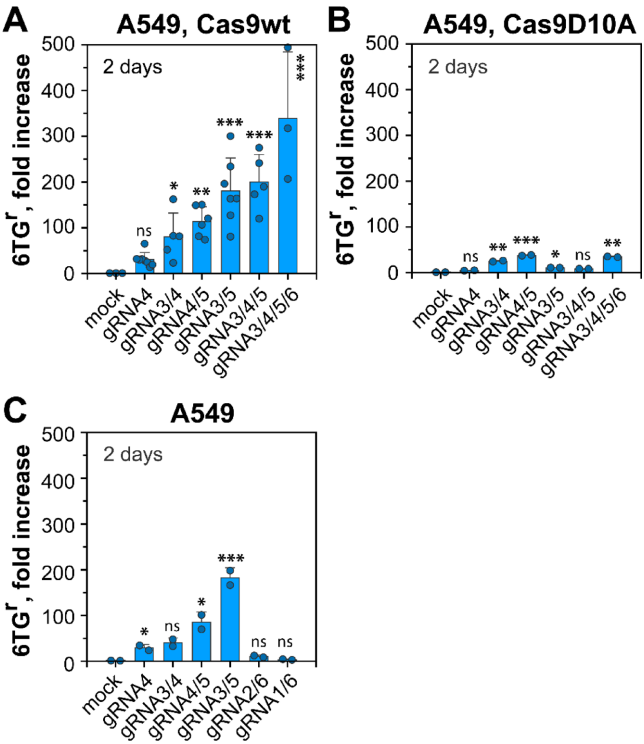
研究人员设计了6条靶向hHPRT基因第3外显子及其侧翼区域的gRNA。通过T7E1酶切实验证实所有gRNA均能有效介导Cas9在预期位点产生DSB。间接免疫荧光显示,转染Cas9与gRNA的细胞中可观察到显著的53BP1和γH2AX焦点形成,而单独转染Cas9的细胞则无此现象,证实了DSB的特异性诱导。
研究结果显示,单个DSB(如gRNA4诱导)可将突变频率提升约55倍。而两个DSB形成的簇可使MF增加约200倍。当DSB簇复杂度增至3个或4个DSB时,MF分别飙升至背景水平的532倍和681倍。这表明DSB簇的突变诱导能力远超单DSB,且随簇内DSB数量增加而增强。值得注意的是,当DSB间距超过约600 bp时(如gRNA1/6组合,相距~1200 bp),协同效应消失,MF甚至低于单DSB,提示DSB的协同效应存在距离依赖性。
为确认DSB本身的关键作用,研究团队使用了仅产生单链断裂(SSB)的Cas9切口酶突变体(Cas9D10A)。结果显示,无论是单SSB还是SSB簇,其诱导HPRT突变的能力均微乎其微,充分证明了DSB的形成是基因失活和突变诱导的核心事件。
通过小干扰RNA(siRNA)敲低DNA-PKcs或使用DNA-PKcs抑制剂NU7441,发现c-NHEJ通路被抑制后,所有类型DSB(单DSB或DSB簇)诱导的MF均显著上升。这表明DNA-PKcs及其介导的c-NHEJ通路在正常情况下是突变的有效"抑制者"。特别有趣的是,在DNA-PKcs基因敲除(A549-PRKDC-/-)细胞中,DSB簇对MF的提升效应随簇复杂度增加而减弱,暗示复杂的DSB簇自身就能部分抑制c-NHEJ活性,因此额外抑制DNA-PKcs产生的增强效应有限。
PARP1基因敲除(A549-PARP1-/-)或使用PARP1抑制剂PJ34,几乎完全消除了DSB簇诱导的突变。同样,抑制与alt-EJ相关的DNA连接酶LIG3(使用抑制剂L67),也显著降低了MF。这表明PARP1和LIG3依赖的alt-EJ是DSB簇诱变的主要通路。即使在DNA-PKcs缺陷的细胞中,PARP1抑制依然能阻断突变诱导,进一步证实alt-EJ的核心作用。
ATM基因敲除(A549-ATM-/-)或使用ATM抑制剂KU55933,降低了DSB簇的突变诱导能力。ATM是DNA损伤应答的关键激酶,参与调控DNA末端切除(resection)。而末端切除是alt-EJ等错误倾向修复通路启动的前提。进一步敲低末端切除关键蛋白CtIP,也显著抑制了MF,但效果不如PARP1抑制彻底,提示可能存在部分不依赖于经典切除的alt-EJ机制。
该研究得出结论:DSB簇是比单DSB更强的诱变因素,其效力随复杂度增加而提升;DSB簇通过空间位阻或染色质结构破坏,内在性地抑制了高保真的c-NHEJ通路;修复"重心"从而转向易错的alt-EJ通路,该过程依赖于PARP1和LIG3活性;ATM激酶通过促进DNA末端切除,在调控这一修复路径转换中扮演重要角色。
此项工作的意义深远。它首次在单基因位点上精准演示了DSB复杂度如何直接决定修复路径的选择和突变结局,为理解高LET辐射的高RBE提供了明确的分子机制解释。所建立的CRISPR/Cas9 DSB簇模型,为未来研究不同染色质环境、细胞周期时相下的DSB簇修复规律提供了强大工具。在应用层面,该研究提示,在利用高LET辐射进行肿瘤治疗时,应密切关注其可能引发的次级突变风险。同时,针对PARP1或alt-EJ通路成分的抑制剂,或许能与高LET放疗产生协同效应,或用于保护正常组织。总之,这项工作将DNA损伤复杂度从抽象概念转化为可精确操作的生物学变量,开创了基因组稳定性研究的新范式。
 生物通微信公众号
生物通微信公众号
 生物通新浪微博
生物通新浪微博
今日动态 |
人才市场 |
新技术专栏 |
中国科学人 |
云展台 |
BioHot |
云讲堂直播 |
会展中心 |
特价专栏 |
技术快讯 |
免费试用
版权所有 生物通
Copyright© eBiotrade.com, All Rights Reserved
联系信箱:
粤ICP备09063491号