
-

����ͨ��
����ץס�����Ƽ�
����������
����ͨ��
����ץס�����Ƽ�
����������
Cell | �����ݿ�����������������������ֲ����ź�ת���»���
�����壺 �� �� С �� ʱ�䣺2024��09��26�� ��Դ��������ѧ�ִ�ũѧԺ
�༭�Ƽ���
���� 2024��9��24�գ�������ѧ�ִ�ũѧԺ�����ݿ�����������������������Cell��������ΪLight-induced remodeling of phytochrome B enables signal transduction by phytochrome-interacting factor���о����ģ���ʾ�˳��������ڴ���phyB���ź�ת���������Ӧ����
̫���ⲻ����ֲ��������������Դ�����ǵ���ֲ�������������̵Ĺؼ��źš�����ɫ����ֲ��ĺ��/Զ������������Ϊֲ��ġ��۾���������Ҫ��ɫ������ȫ������仯����Ƶ����������Դ��������ά��ȫ����ʳ��ȫ��һ������ս����ȥ��һ�����ͣ�ũ��������IJ�����ߵ�����Խ��Խ����ֲ��������Ʒ�֡��������������һ�����Ҳ���������������������ܼ���ֲ����Ʒ�֡�������ֲ������ֲ��ĸй���������ܸ߲���״����ֲ�����ɫ�ص���ϢϢ��أ�����ֲ�����ɫ����Ӧ�Լ����ݹ��źŵĻ��ƽ������ڸ���������ֲ���ܼ��Ը��ӻ�������Ӧ�ԣ��Ӷ�Ϊ��ʳ��ȫ�ı����������ס�
�ߵ�ֲ����Ҫ�����Թ���ɫ��A��phyA������ɫ��B��phyB��Ϊ�������������ɫ�أ�����phyB�ǽ鵼��������Ӧ����Ҫ������塣����ɫ��ͨ����ɫ��P��B�ں������̬��Pr����̬���Լ�Զ�������̬��Pfr������̬��֮����п���ת�䡣���Ͻ�phyB���⼤�����ֱ����һ�����ɫ�ػ������ӣ�phytochrome-interacting factor��PIF)���������ݹ��źŲ��������λ������ٽ�����̬���ɡ����phyB��PIFs������ֲ����Ӧ��Χ����Ĺؼ��ź�ģ�顣���Ͻ���8��PIF��Ա��PIF1-8��������������Ҫ�ṹ��N��Ϊ���phyB-Pfr�ļ���̬�����Active-PHYB Binding motif��APB������C��Ϊ���DNA��bHLH���ۻ��ṹ���ںڰ������£�phyB-Pr��λ��ϸ�����У�PIF1/3/4/5�ں��������������簵��̬���ɵķ������̣��������쳤����Ҷ�պϣ���һ�������֪��⣬phyB-Pfr��˽�������������PIFs�������������쳤�ʹٽ���Ҷչ������ά��ֲ���������̬������2022������Vierstra�Ŷ���Nature���Ľ�ʾ��phyB-Pr�Ľṹ����phyB-Pfr����ʶ��PIF�Ľṹ����ѧ����������ػ����Բ������
2024��9��24�գ�������ѧ�ִ�ũѧԺ�������������������������������Cell��������ΪLight-induced remodeling of phytochrome B enables signal transduction by phytochrome-interacting factor���о����ģ���ʾ�˳��������ڴ���phyB���ź�ת���������Ӧ���ơ�

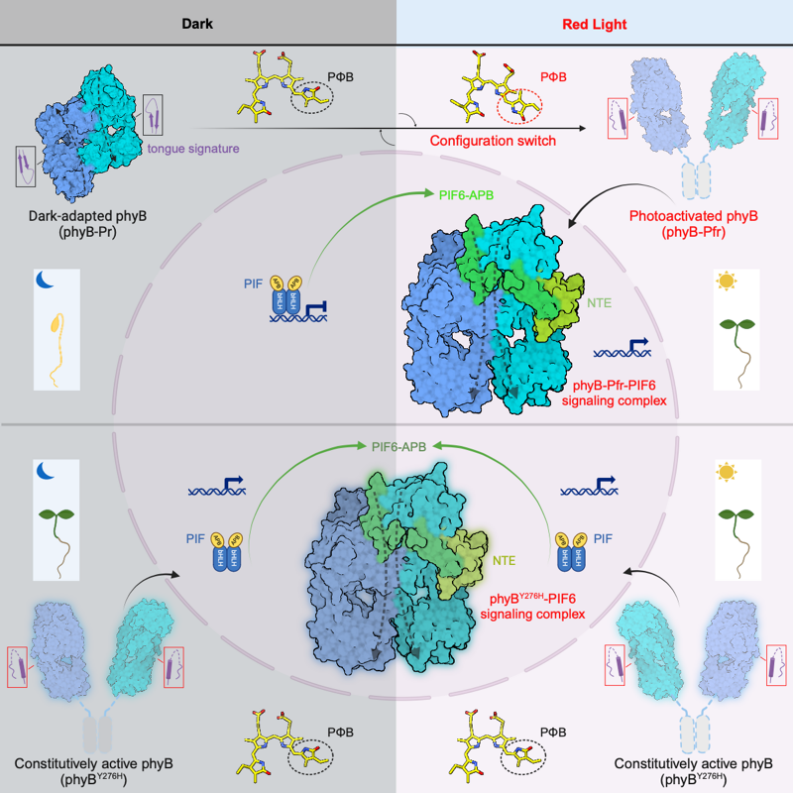
���о�������ģʽֲ�����Ͻ�⼤��̬phyB-Pfr�Լ��������ڹ������ͼ���ͻ����phyBY276H�ֱ��������źŷ���PIF6�ĸ�����߷ֱ����䶳�羵�ṹ���ֱ�������Ϊ3.1 ?��3.2 ?�������ڷ��ӽṹ���������ͻ���嵰�������ʲⶨ�������Լ�����������ʵ��������Լ�ת����ֲ��ı��ͷ�������ʾ�˹⼤��phyB��Prת��ΪPfr����ϸ���ӻ��ƣ��������phyB��PIF֮�䡰�յ�-���ϡ��������ģ�ͣ����ֲ�����ɫ���źŴ��������о���һ���ؼ��հס�
����ȫ��PIFs���״��ڴ�������ṹ��������������Լ���װphyB-PIFȫ��������������൱������ѡ����ǵ�ϵ�����ױ�����PIFs��APB motif��N��100�������ᣩ���Խ鵼����phyB�����ڼ����⻥�����о���ѡȡ�˽��������ǿ��PIF6-APB��PIF6-100����N��100�������ᣩ����phyB-PIF�ĸ�������װ���Ʊ��䶳��Ʒ�����ճɹ������õ���phyB-Pfr-PIF6�Լ�ȱʧHKRD�ṹ����N��908�������ᣩ��phyBY276H-908-PIF6������ĸ߷ֱ����䶳�羵�ṹ���ṹ��������phyB-Pfr�ڹ⼤��֮�������ģ�ṹ���ţ�����Ӧģ�飨photosensory module, PSM����Pr״̬����ͷ��β��ת��ΪPfr״̬����ͷ��ͷ�������壬��PIF6-APB��������phyB-Pfr����������һ�࣬���phyB-PIF6�����塣���⣬phyBY276H-908-PIF6������ṹ��phyB-Pfr-PIF6������ȫһ�£�ͼ1����
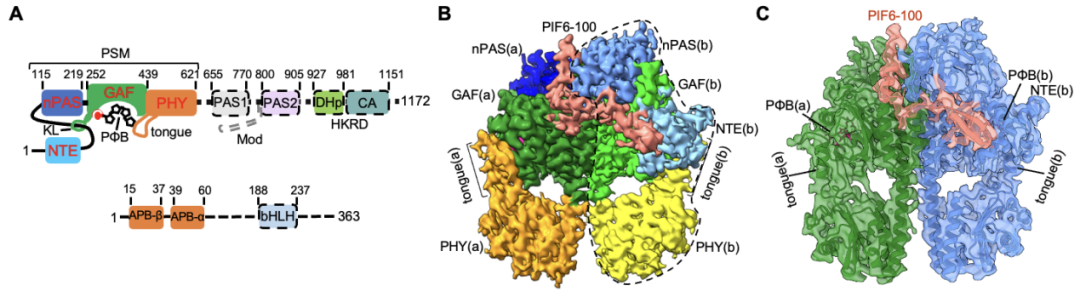
ͼ1. phyB-Pfr-PIF6��B����phyBY276H-PIF6��C����Ϊ���Գ�������
Ϊ�˲����⼤������phyB�Ľṹ���ţ�����ϸ�±ȶ���֮ǰ���ױ�����phyB-Pr�����о���õ�phyB-Pfr�ṹ�����ַ�ɫ��P��B���������պ�����D����ת��180?�����ҺͿڴ��е�ϵ�а��������½����������磨ͼ2A�������յ���PHY�ṹ�������以������״ͻ���ṹ��������Ƭ�㵽�������Ĺ���ת�䣨ͼ2A���ң����ṹ����������˿����S584�����ȶ���������ʽ����״�ṹ�dz���Ҫ��ͼ2A���ң�����һ�������������Լ�ת����ֲ����ͷ���֤����S584����άϵphyB-Pfr����״̬��phyB�ź�ͨ·������Ҫ��ͼ2B����

ͼ2. ���յ�P��B���ӣ�A����PHY�ṹ����״ͻ���ṹ��A���ң��Ĺ���ת�估������֤��B��
��Pr״̬�£�phyB��Cĩ�˽ṹ��PAS2�Լ�HKRD��Nĩ�˽ṹ��GAF��PHY������άϵ��ͷ��β�������幹����phyB-Pfr�У���PHY�ṹ�����״ͻ���ṹת��Ϊ��������ֱ�Ӻ�Pr״̬���PAS2�ṹ�����ռ��ͻ��ͼ3A����ͼ3B�����Ӷ��ƻ�Pr״̬��PAS2��PHY�ṹ��֮��Ĺ㷺����������һ���ƻ�HKRD��PHY�ṹ��ͼ3A����ͼ3B���У����Լ�HKRD��GAF�ṹ��ͼ3A���ң�ͼ3B���ң�֮��ķ����ڻ�������һϵ�еĹ���仯��ȫ����Pr״̬�µ���ͷ��β���ṹ�����ռ���phyB�����⣬ȱʧHKRD�ṹ���phyB�ض��嵰�ף�phyB-908������N��908�������ᣩ�ڹ⼤��֮�������ڵ�����ʽ���ڣ�ͼ3C������һ����֤��phyB-Pr�ڹ⼤���ԭ����N�˺�C��ģ�黥��άϵ����ͷ��β�������屻���ƣ������γ�phyB-Pfr��
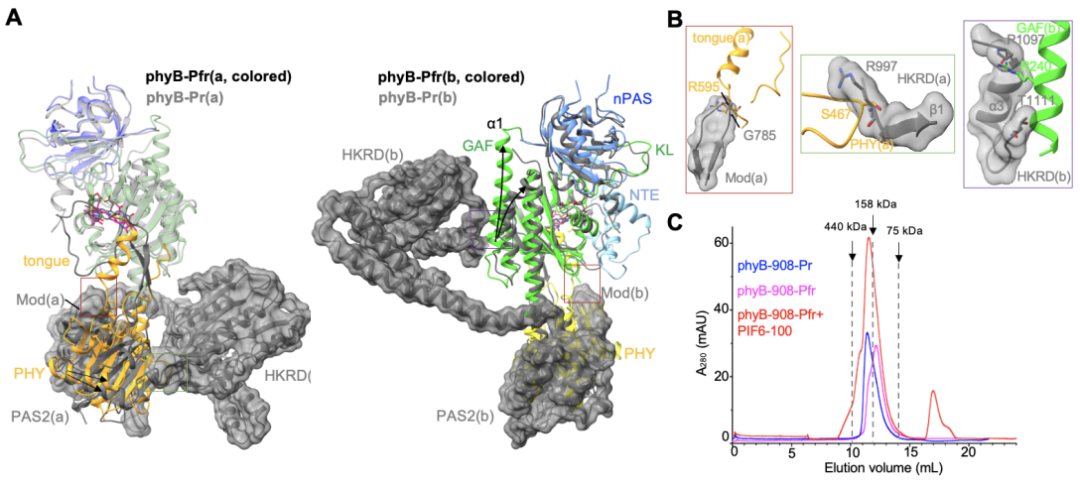
ͼ3. PHY �ṹ����״ͻ���ṹ�Ĺ���ת������phyB-Pr�ع��γ�phyB-Pfr��A��B����������֤��C��
�⼤����phyB-Pfr�ֱ�������Nĩ�˵�NTE�ṹ���Լ�������Nĩ�˽ṹ���γɵ���ͷ��ͷ�������壬��ʶ��ͽ��PIF6-APB��Nĩ�˺�Cĩ�ˣ�ͼ4A����PIF6-APB��Nĩ��ͨ���γ�һ�������нṹ��PIF6-APB-������phyB-Pfr��NTE�ṹ�����㷺������ͼ4B������phyB-Pfr������ṹ����һ����phyB�����߱��ȶ��ṹ��NTE������NTE���ȶ��ṹ����PIF6-APB�Ľ�����յ��γɵġ����⣬PIF6-APB��Cĩ��ͨ���γ�һ���������ṹ��PIF6-APB-����ͬʱ������phyB-Pfr��Nĩ�˽ṹ������㷺������ͼ4C����ӡ֤��PIF6-100�ٽ���phyB-908-Pfr�Ķ��ۻ���ͼ3C�������߽�һ��ͨ��ϵ������������ʵ����������ת����ֲ��ı�����ȷ��NTE�ṹ���Լ�Nĩ�˽ṹ��鵼��phyB-Pfr���ۻ�����phyB-PIF��������Ҫ�ԡ�
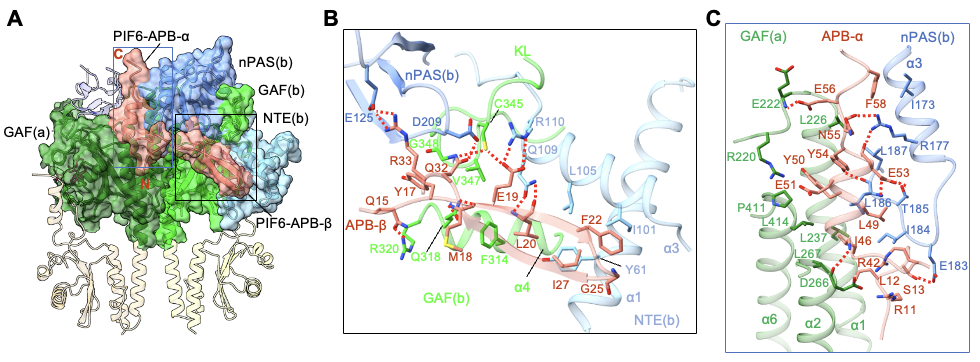
ͼ4. phyB-Pfr��Nĩ�˽ṹ���PIF-APB��������ʶ��A���Լ�NTE�ṹ��������ʶ��PIF-APB��Nĩ�������нṹ��B����phyB-Pfr������������ʶ��PIF-APB��Cĩ���������ṹ��C��
����phyB-Pfr�Ľṹ�ȶԱ���phyB-PIF6������ṹ�е�phyB-Pfr�γ��˲��ԳƵĶ����壨ͼ5A������Щ���Գ��Ե���phyB-Pfr���������һ�����û���㹻�ռ���PIF6-APB-���Լ�PIF6-APB-���γɺ��ʵĻ�����ͼ5B�������߽�һ��ͨ��pull-down�Լ�Co-IP assay��������Һ״̬��phyB��PIF-APB�Լ�PIF-FL�Ļ���ģʽ���������ݱ���PIF3��PIF6��APB motif�Լ�PIF1��PIF3��ȫ���������Ժ�phyB-Pfr�������γ�Ħ����Ϊ1:2��2:4���ĸ����
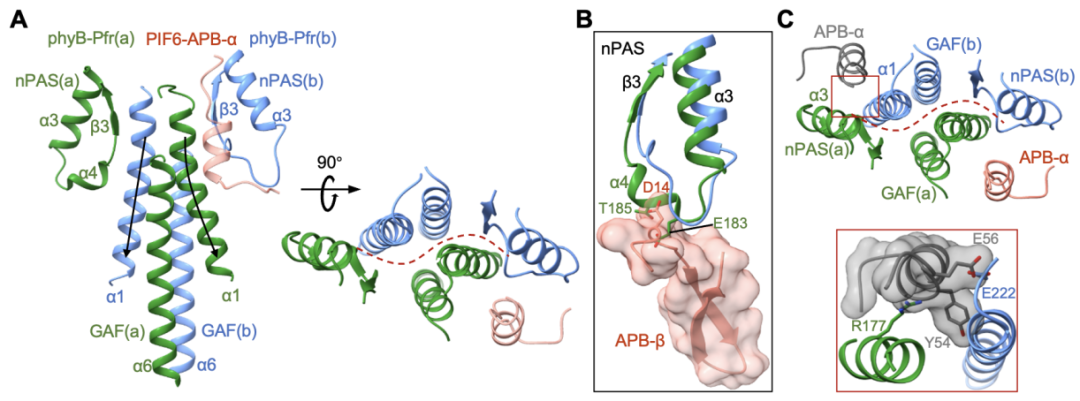
ͼ5. ���ԳƵ�phyB-Pfr������ֻ�ܽ��һ��PIF6-APB����
�ܵ���˵�������о���ʾ�ˡ�ͷ��ͷ������̬phyB������ʶ����PIF6-APB�ĸ�����ṹ������˹���ɫ���о��й�����յ�phyB�乹���ת��PIF�źŵĹؼ����⣨ͼ6����Ϊ����й���״�ĸ����Լ�phyB��ع�ػ������ġ����Ŵ�ѧ�����߿����ṩ�˷���ˮƽ�ľ�ϸͼֽ��
ͼ6. ��⼤��phyB��ת��PIF�źŵĽṹ����ģ��
������ѧ�ִ�ũѧԺ��������ѧ�ִ�ũҵ�о�Ժ�������о�Ա������������Ϊ�����ĵ�ͨѶ���ߡ�������ѧǰ�ؽ���ѧ���о�Ժ��ʿ�о���������������Ƽ��������ѧ�ִ�ũҵ�о�Ժ�����������ķ����Եϵ����Լ�������ѧ�ִ�ũҵ�о�Ժ��������ʿΪ���Ĺ�ͬ��һ���ߡ�������ѧ�ִ�ũҵ�о�Ժ�ԬB��ʿ���ٳ���ʿ������ʦ������й���ѧԺ�Ŵ��뷢������ѧ�о�����ʿ�о��������Ա��о�Ҳ��������Ҫ���ס��䶳��Ʒ�Ʊ�����Ʒɸѡ�������ռ����о�Ժ�����۽ṹ�о�ƽ̨��ɡ����о��õ��˹�����Ȼ��ѧ���𡢹����ص��з���Ŀ��ɽ��ʡ�ص��з���Ŀ��ɽ��ʡ�Ƽ����»����п�Э�����˲��оٹ��̡�ɽ��ʡ̩ɽ����ѧ����Ŀ��������ѧ�ִ�ũҵ�о�Ժ��Ϋ���ִ�ũҵɽ��ʡʵ���ҡ�С������ȫ���ص�ʵ���ҡ�������ѧ��������ֲ������о������ص�ʵ������ؾ��ѵ�������
�������ӣ�https://www.cell.com/cell/fulltext/S0092-8674(24)01023-7
ר�ҹ۵�
���� Ժʿ���й�ũҵ��ѧԺ�����ѧ�о���
������ֲ��������������Դ����ũ����������ž��������á���ũҵ�����У����������ֲ�ܶ�����������Ҫ;����Ȼ���������ܶ����ӣ�����ֲ��֮���������Ӿ磬���ֲ�꾥�����ߡ�����������������С��Ϳ����Լ�����ͻ�����⡣��ˣ�����ֲ���֪���յķ��ӻ���������������Ӧ�����������ԵȾ���ʮ����Ҫ�����塣����ɫ����ֲ��ĺ�⡢Զ������壬��Ӱ��������Ӧ���ձ仯�Ĺؼ����ܵ��ס���1960���״���������ɫ������������������������о���Աһֱ�������ɫ�ظ�֪���/Զ����źŵĻ��ơ�����������80���ĩ���о���Ա��ֲ�����ɫ��phyA�ĵ羵��Ƭ�оͿ�������Y�����εĵ��������������ڹ���ɫ�����ڶ�ṹ������嵰�ף�ֲ����Դ�����ϵͣ�������������ѣ��о���Ա��Ҫͨ�����������õ�������Դ����ɫ�ؽ��нṹ������ֲ�����ɫ�صĽṹ�о����ͣ�͡�ֱ��2014�꣬PNAS��־�����˻�̬phyB���Ӧģ��ض���ľ���ṹ��2022�������������������Լ��й��Ķ�ҵ�λ��̱�����ȫ��phyB��phyA��̬�Ľṹ���Ӷ������˹���ɫ�ػ�̬�ṹ���Ե���ʶ����ֲ�����ɫ�ؼ���̬�ṹ�����ź�ԭ��ת���Ļ����Բ�����������˻��ڼ���̬phyB�ṹ�Ĺ��ź�ת�������о���
�����о��״α����˼���ֲ̬�����ɫ��phyB��ϱ�־�������źŷ���PIF�ĸ�����ṹ����ʾ�˹㷺�о��IJ���������յ�����ͼ���ͻ����phyBY276H���PIF�ĸ�����ṹ������϶����о������������˹�������ļ���̬phyB�ṹ�����Լ�����ɫ�ص��ͷ�ɫ�ŷ���֮���Э���Ķ�̬�仯���ơ������о���ֻش��˳��������о���Ա��ע��phyB�й⼤���������˼���ֲ̬�����ɫ�ؽṹ�о��Ŀհף�������ѧ�����о����ش�ͻ�ơ�ͬʱ������ɫ�ؼ�����ƵĽ���Ϊ������ֲ�����Ӧ�����˻������Զ���������ס�С��ˮ������Ҫ����ĸй���״���������ܸ߲�Ʒ�������ش�
������ ����������ũҵ��ѧ��
��������ֲ������������ﵥ������Ч������ʩ�������Ǿ������������ԵĹؼ�����֮һ��Ȼ������ֲ�����շ�ֲ���������Ӧ������ֲ��ͽ�������ѱ�ϸ����ϵ�������������ӵ������յȲ����������Ϊֲ����Ӧ���/Զ����źŵ���Ҫ������壬����ɫ��phyB����ֲ��Ӧ��������仯����Ӧ����������ߡ�Ҷ�нǡ�����ʱ��ȣ�������Ҫ���Ӷ�Ӱ����������͡����Ч�ʺͲ�����phyB����⼤���ͨ����PIF����ֱ�ӻ������ݹ��źţ�����������ά�ֹ���̬���ɵķ������̡�Ȼ����1998�귢��PIF�Ǽ���̬����ɫ�ؽ�ϵ�����������Ȼ�����˲���Ӱ��phyB-PIF�����Ĺؼ�λ�㣬������̬phyB��PIF�������Ľṹ�����͵��ػ���Ŀǰ��Ȼδ֪�������о�ͨ��������ģʽֲ�����Ͻ�⼤��̬phyB-Pfr�Լ��������ڹ������ͼ���ͻ����phyBY276H�ֱ��������źŷ���PIF6�ĸ�����߷ֱ����䶳�羵�ṹ����ʾ��phyB��̬��Pr���ڹ⼤�������Ӧģ�飨PSM����Pr״̬����ͷ��β��ת��ΪPfr״̬����ͷ��ͷ�������塣����phyB-Pfr������IJ��Գ��ԣ��������һ���ܹ����PIF���γ�phyB-Pfr-PIF�������塣���߽�һ�����ͻ���嵰�������ʲⶨ�������Լ�����������ʵ��������Լ�ת����ֲ��ı��ͷ�������֤��˿����S584��NTE�ṹ���Լ�Nĩ�˽ṹ����ڽ鵼phyB-Pfr���ۻ���phyB-PIF������phyB�ź�ͨ·����Ҫ�ԡ����о��״ν�ʾ��phyB�⼤�������źŴ��ݣ���PIF�������Ľṹ����ѧ�������ǽ�����ֲ�����ɫ���źŴ����о�����ȡ�õ���һ���ش�ͻ�ơ��о��ɹ���Ϊ�����������ܡ����������߹�Ч������Ʒ���ṩ��Ҫ����ָ���ͻ�����Դ��
����ͨ�Ź��ں�
֪����ҵ��Ƹ

