
-

生物通官微
陪你抓住生命科技
跳动的脉搏
生物通官微
陪你抓住生命科技
跳动的脉搏
张勇和王皓毅团队合作探索DNA转座子多样性并拓展基因工程工具箱
【字体: 大 中 小 】 时间:2024年06月07日 来源:中国科学院动物研究所
编辑推荐:
2024年6月5日,中国科学院动物研究所张勇和王皓毅研究组在Cell杂志上在线发表题为“Heterologous survey of 130 DNA transposons in human cells highlights their functional divergence and expands the genome...
自1948年Barbara McClintock首次报道转座子以来1,这类跳跃遗传元件对宿主演化的重要意义逐渐被揭示2-5。其中,DNA转座子作为主要类型之一,长期以来备受关注。然而,由于以往多为个案研究,DNA转座子活性的决定因素与进化模式的一般规律尚不清晰。不仅如此,尽管DNA转座子可作为基因工程工具用于插入诱变或转基因载体6,7,但目前只有Sleeping Beauty(SB)等少数几种转座子得到了开发和广泛应用。因此,系统挖掘具有不同功能特点的转座子工具亟待开展8。
2024年6月5日,中国科学院动物研究所张勇和王皓毅研究组在Cell杂志上在线发表题为“Heterologous survey of 130 DNA transposons in human cells highlights their functional divergence and expands the genome engineering toolbox”的研究论文。该研究筛选到40个在人源细胞中具有转座活性的新型DNA转座子,系统解析了DNA转座子的活性相关因素和进化动态。这些具有不同功能特点的转座子拓展了相关基因工程工具箱;尤为重要的是转座子Mariner2_AG(MAG)在CAR-T免疫治疗实验中展示出高活性,具有临床应用价值。
具体而言,研究团队首先建立了遗传多样性数据的挖掘模式(图1A),基于生物信息分析从102个无脊椎和脊椎动物基因组中预测了130个潜在活跃的DNA转座子;人源细胞中的高通量筛选实验进一步验证了40个转座子具有异源转座活性(图1B)。其中,来自蚊子的Mariner2_AG、来自洞穴鱼的Tc1-2_St等转座子活性超越了经过大量优化的SB100X。
这一结果将哺乳动物中活跃转座子载体的数目从20提升至了60,大幅拓展了其进化多样性(图2)。研究团队通过比较活跃与不活跃的转座子,发现高活性DNA转座子富集在Tc1/mariner超家族;这与该超家族转座子频繁发生水平基因转移的特点相吻合。在转座子的来源物种、家族、年龄和完整拷贝数等各类进化信息中,拷贝数最能准确预测其在人类细胞中的活性有无。
研究者探究了影响转座活性的序列特点。在Tc1/mariner超家族中,转座子末端的反向重复(TIR)如果相对较长并其内部包含正向重复(IR-DR),那么该转座子更有可能有活性(图2、3A),且其活性也较高(图3B)。除了非编码区特征外,研究者也在转座酶编码区证实了Tc1家族和hAT超家族中对转座活性具有重要影响的已知功能性位点的绝对保守性9,10,同时还报道了更多新的功能氨基酸位点(图3C)。
考虑到其作为基因工程工具的潜在可能,研究者进一步探究了新型转座子的整合偏好性和基因承载能力等功能属性。来源于不同超家族的高活性转座子展示出三种整合偏好模式(图4):一是接近于随机的整合模式,如Tc1家族成员;二是偏好于异染色质区域,如piggyBac-1_AMi转座子;三是偏好于转录活跃区域,如hAT超家族成员。这些DNA转座子的整合偏好性差异代表其在不同领域的应用潜力,例如第三种转座子可用于增强子或基因定位分析。基因承载能力分析则显示出活性最高的Mariner2_AG转座子在该方面的明显优势:其在承载10kb的外源基因时仍可表现出高达30%的转座效率。
研究团队最后对MAG转座子用于CAR-T免疫治疗的能力进行了初步探索,揭示了其作为基因治疗载体的优越性(图5)。相比于常用的慢病毒和SB100X载体(特别是前者),MAG转座子制备的CAR-T细胞在人 Burkitt's 淋巴瘤细胞系Raji构建的恶性血液瘤模型和人肺鳞癌细胞系NCI-H226构建的实体瘤模型中展现了更佳的治疗效果与更长的存活时间。
综上所述,该研究基于自然界的动物遗传多样性开展了迄今为止最大规模的DNA转座子活性筛选,获得了目前最大的活跃DNA转座子载体数据集。这不仅揭示了与转座活性相关的进化因素和功能特征,还扩展了基于DNA转座子的遗传工具箱。
该研究由中国科学院动物研究所、中国科学院大学、北京中医药大学、四川大学华西医院等机构合作完成。动物研究所博士生张童童、副研究员谭生军、助理研究员唐娜、博士生李源清为该研究共同第一作者,动物研究所张勇研究员和王皓毅研究员为共同通讯作者。张晨泽、孙晶、郭言言、高晖、蔡瑜佳、孙文、王晨鑫、傅良政、马慧静、吴亚超、胡晓璇、张雪纯、Peter Gee、闫卫华、赵亚辉、陈强、郭宝成等合作者在文章写作、实验和计算分析等方面提供了大力支持。该研究得到了自然科学基金(32325014)、农业农村部(2023ZD0407401)、重点研发计划(2019YFA0802600,2019YFA0110000)、科学院(ZDBS-LY-SM005,XDA16010503)等项目资助。
论文链接:https://doi.org/10.1016/j.cell.2024.05.007
代表性参考文献:
1.Mcclintock,B. (1948). Mutable Loci in Maize. In (Carnegie Institute of Washington Year Book),pp. 155-169.
2.Almeida,M.V.,Vernaz,G.,Putman,A.L.,and Miska,E.A. (2022). Taming transposable elements in vertebrates: from epigenetic silencing to domestication. Trends in Genetics 38,529-553. 10.1016/j.tig.2022.02.009.
3.Moon,S.,Cassani,M.,Lin,Y.A.,Wang,L.,Dou,K.,and Zhang,Z.Z. (2018). A robust transposon-endogenizing response from germline stem cells. Developmental cell 47,660-671. e663. 10.1016/j.devcel.2018.10.011.
4.Wells,J.N.,and Feschotte,C. (2020). A field guide to eukaryotic transposable elements. Annual review of genetics 54,539-561. 10.1146/annurev-genet-040620-022145. Epub 2020 Sep 21.
5.Lohe,A.R.,Moriyama,E.N.,Lidholm,D.-A.,and Hartl,D.L. (1995). Horizontal transmission,vertical inactivation,and stochastic loss of mariner-like transposable elements. Molecular biology and evolution 12,62-72. 10.1093/oxfordjournals.molbev.a040191.
6.Rubin,G.M.,and Spradling,A.C. (1982). Genetic transformation of Drosophila with transposable element vectors. Science 218,348-353. DOI: 10.1126/science.6289436.
7.Zhao,D.,Ferguson,A.,and Jiang,N. (2015). Transposition of a rice Mutator-like element in the yeast Saccharomyces cerevisiae. The Plant Cell 27,132-148. 10.1105/tpc.114.128488.
8.Kebriaei,P.,Izsvák,Z.,Narayanavari,S.A.,Singh,H.,and Ivics,Z. (2017). Gene therapy with the sleeping beauty transposon system. Trends in Genetics 33,852-870. 10.1016/j.tig.2017.08.008.
9.Plasterk,R.H.,Izsvák,Z.,and Ivics,Z. (1999). Resident aliens: the Tc1/mariner superfamily of transposable elements. Trends in genetics 15,326-332. 10.1016/s0168-9525(99)01777-1.
10.Yuan,Y.W.,and Wessler,S.R. (2011). The catalytic domain of all eukaryotic cut-and-paste transposase superfamilies. Proc Natl Acad Sci U S A 108,7884-7889. 10.1073/pnas.1104208108.
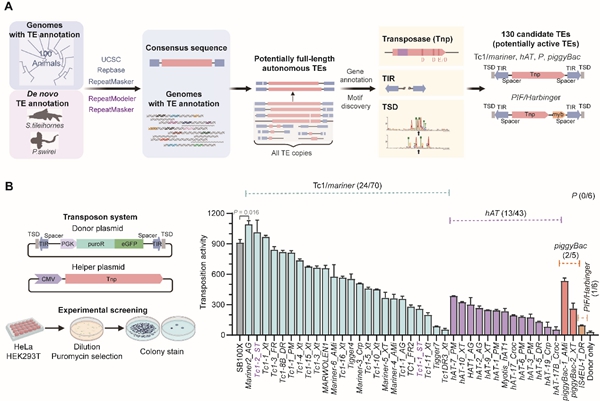
图1. 系统筛选活跃DNA转座子。A)预测活跃DNA转座子的生物信息学分析流程。B)实验验证体系及40个活跃DNA转座子的转座活性。
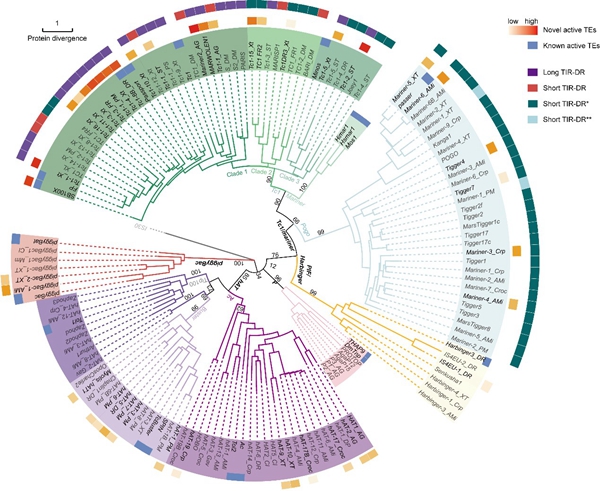
图2. 130个新型DNA转座子和20个已知活跃DNA转座子的进化树。
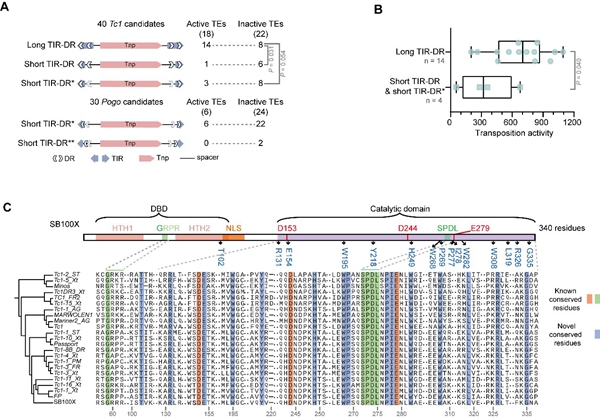
图3. 活跃转座子的固有属性。A)Tc1/mariner超家族转座子的四种TIR-DR结构。B)Tc1家族中,具有长TIR和短TIR的活跃转座子的转座活性分布情况。C)Tc1家族活跃转座酶的保守氨基酸位点。
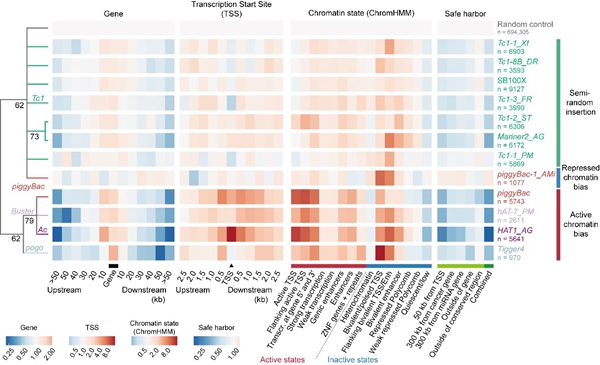
图4. 不同家族高活性转座子的插入偏好。
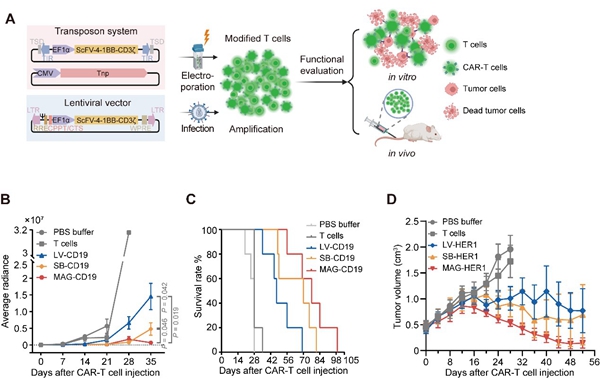
图5. MAG转座子制备的CAR-T细胞功能评估。A)CAR-T细胞制备及功能评估流程。B)接种人 Burkitt's 淋巴瘤细胞系Raji的小鼠在接受不同处理后的荷瘤情况。C)接种人 Burkitt's 淋巴瘤细胞系Raji的小鼠在接受不同处理后的生存曲线。D)接种人肺鳞癌细胞系NCI-H226的小鼠在接受不同处理后的肿瘤体积变化。

