
-

����ͨ��
����ץס�����Ƽ�
����������
����ͨ��
����ץס�����Ƽ�
����������
����ֲ����������Ա��������߽�ʾ����ֲ������ݻ���ʷ�ؼ�ģʽ
�����壺 �� �� С �� ʱ�䣺2024��12��27�� ��Դ���й���ѧԺ����ֲ���о���
�༭�Ƽ���
�������о��ɹ��� Convergent patterns of karyotype evolution underlying karyotype uniformity in conifers Ϊ�ⷢ���� Wiley���¿��Ż�ȡ�콢�ڿ� Advanced Science ��
�������ݻ��Ĺ����У����͵��ȶ��Ժͱ�������ì�ܵ��������档һ���棬���͵��ȶ��Զ��Ŵ���Ϣ����������������Ҫ����һ���棬���͵ı����Զ������γɺͶ�����Ҳ����Ҫ���á���ͬ�ڱ���ֲ������ĺ��ͣ�����ֲ��ر����ɰ��ࣩ���ֳ��൱�ߵĺ���һ���ԡ�����һ����������γɺ�ά�ֵģ�һֱ��δ��֮�����й���ѧԺ����ֲ���о��������й���ѧԺֲ���о����������Ĭ�´�ѧ�о��Ŷӣ�ͨ���Ƚϻ�����������ؽ�������ֲ������ݻ���ʷ����ʾ���ɰ�������ݻ��Ĺؼ�ģʽ��Ϊ����ֲ������ֶ������ݻ��뱣���ṩ���µ��ӽ���
�о��Ŷ�ͨ����16������ֲ��Ⱦɫ��ˮƽ�Ļ����鹲���Է����������ɿơ��ؿơ��춹ɼ�ơ����ӡ����������������ʾ��һ��һ�Ĺ����ԣ�������Щ���ֶ�δ����������ϵ���еĹŶ���¼���ͼ1���������ų��˹Ŷ��������ֲ����Ҫ��֧������������⣩���͵�Ӱ�졣������ģʽ��һ�������ɰ��������䷢����Ƶ���ķ�ͬԴȾɫ��֮������λ(reciprocal translocations,?RTs)����Щ��λ��Ⱦɫ���Ϊ��λ����ϵ�ͨ����λ����˿��������ͼ2����������Ϊ��˿���C��˿�����λ(centromeric�Ccentromeric reciprocal translocations,?CRTs)�����⣬�о���Ա�ڰ�Ŀ�������з�������һ�����λ����ϵ�ֱ�λ��һ��Ⱦɫ�����˿����������һ��Ⱦɫ��Ķ����������������Ϊ�����C��˿�����λ(telomeric�Ccentromeric?reciprocal translocations,?TRTs)����һ�����о�ȷ�ϣ�CRTs���ɰ�������ݻ�������Ҫģʽ����TRTs���ڰ�Ŀֲ���еĵ����챶��(descending dysploidy)�з����˹ؼ����á�
����CRTs����Ƶ����������������ֲ������ݻ�·�����پ���ս���о��Ŷ���ͼ�۵ķ���(graph-based method)����CRTs�ķ���·����ͼ2��������һ���ؽ�������ֲ��ӹ�ͬ���Ⱥ���(proto-gymnosperm karyotype,PGK��n?= 12)���ִ�����(n?= 11�C12)����ϸ�ݻ�·����ͼ3�����ؽ������ʾ���ɿ��У��ȷ�����5�ι���CRTs��֮�������з�����һ������CRT��chr1��chr3֮�䣩�����ɼ���������Ҳ����һ������CRT��chr5��chr11֮�䣩����ЩCRTs������7��Ⱦɫ���������ϣ���ʣ��5��Ⱦɫ����������״̬��ͼ3A�����ڰ�Ŀ�������Ϊ�������������е�Ⱦɫ�������PGK��������������ϣ�ģ���������CRTs�ܿ����Ѿ����ͣ�ͼ4����������ƶϲ��ַ�֧��CRTs������ȷ�д�����ͼ3A�������⣬�ƶϰ�Ŀ��ͬ����һ��TRT�¼���������һ��ֻ��һ������Ⱦɫ��۵Ķ���˿��Ⱦɫ�壬֮���ڲ�ͬ��֧�������˲�ͬ���ݻ�·����ͼ3B�����ڰؿ��У�����Ⱦɫ��Ķ̱ۣ���������ͨ������5��CRTs���ݸ���PGK-6S��PGK 6��Ⱦɫ��̱ۣ���֮����ͨ��һ��TRT��PGK-6S����˿������PGK-5S�Ķ����˷����ںϣ��γɵ�������������һ�����̵�Ⱦɫ�壬�����ն�ʧ�����°ؿ�����Ⱦɫ����Ŀ���͵�n?= 11��ͼ3B�������Ƶĵݽ��챶��Ҳ������������鼻����飨n?= 11���С��ں춹ɼ��n?= 12���У���������˿��Ⱦɫ����ȻҲ������һ��CRT�������ɴ�������ͼ3B������֮���ڰ�Ŀ�У�����һ��TRT�ı���Ⱦɫ���ṹ����̬��֮��ͬ��ϵ���ؿƺ���鼣�����������TRTs�ֽ�����Ⱦɫ����Ŀ��
�о���Ա��������CRTs��TRTs���ܵķ������ƺ�DZ��Ӱ�졣CRTsƵ����������˿������������Ϊ�ɰ������ձ���ڵ�����������(Rabl configuration)����˿���ۼ���һ�𣬴ٽ�����˿���������ú���λ����˿������������ظ�����Ҳ�ܹ��ٽ��ǵ�λͬԴ����(non-allelic homologous recombination,?NAHR)�鵼�����λ��ֵ��ע����ǣ�����������CRTs����������ͬһ��˿������IJ�ͬλ�á���TRTs����żȻ�����ڼ�������ʱ�ڣ����������ṹ(telomere bouquet)���γɿ��ܴٽ��˶����C��˿���Ļ�������λ����TRT�IJ�������һ�����̵�Ⱦɫ��ʱ������Ⱦɫ����Ϊ�����б����������ܶ�ʧ�����µݽ��챶�ԡ����뱻��ֲ���е�Ⱦɫ��ĩ���ں�(end-to-end fusion)���ƣ�������ֲ���챶�Եķ�������ʮ�ֶ�������������������������TRT�Ļ��ơ�����ֲ�ﳬ����Ⱦɫ��������������������ķ��������⣬CRTs��TRTs�ܹ�������ͬ�ĺ��͡�������ֳ���룬���ܴٽ�������ֲ��������γɺͶ�������
���о��������������Ƕ�����ֲ������ݻ�����ʶ�����Ҷ�ֲ��Ⱦɫ���ݻ��о�������Ҫ�ο���ֵ�����о��ɹ���Convergent patterns of karyotype evolution underlying karyotype uniformity in conifersΪ�ⷢ����Wiley���¿��Ż�ȡ�콢�ڿ�Advanced Science�����й���ѧԺ����ֲ���о������ʸٲ�ʿ�о���Ϊ���ĵ�һ���ߣ�����ֲ�����������о�Ա���������о�Ա�������Ĭ�´�ѧ��ΰ��ʿ���й���ѧԺֲ���о�����Сȫ�о�ԱΪ��ͬͨѶ���ߡ���Ĭ�´�ѧ���Բ�ʿ������ֲ�����к��˲�ʿ�о��������˶ʿ�о������ѱ�ҵ���������Ÿ��о�Ա�����˶ʿ�о�����ɽ��ʡũ��Ժ�ֿ������о�Ա�����˱���������о��õ��˹����ص��з��ƻ�(2022YFF1301700)������ֲ����ս���ȵ��ƻ� (KIBXD202401)������ʡ�ص��з��ƻ�(202403AC100028)����Ŀ�Ĺ�֧ͬ�֡�
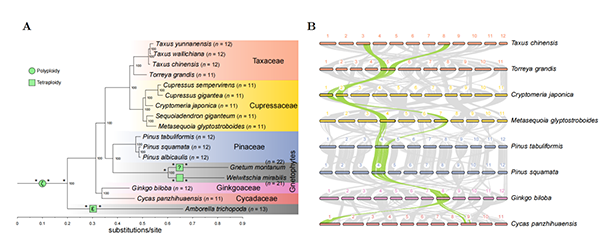
ͼ1. ����ֲ�ﺱ�����Ŷ����WGD���¼����ɿơ��ؿơ��춹ɼ�ơ����ӡ��������������ʾһ��һ�Ĵ��ģ�����ԣ�B����������Щ���ֶ�δ����������ϵ���е�WGD�¼���A����ͬʱ��������֤��Ҳȷ����ǰ�˱����İ�����WGD��ȷ��Ϊ�ı�����������ֲ�ﹲ��WGD����-WGD��ȷ��Ϊ�ı�����������ֲ�ﹲ��WGD����-WGD��δ֪���ԣ���Ҳ��һ��֤�ݱ���������Ҳ������WGD�����ɣ�����Щ���ϵ�WGD�����˺����Ķ�������������Ⱦɫ�����ţ����������������Ĺ�����ʮ�����飬��Ӧ�ĺ����ݻ�·���������ݣ�A����*������

ͼ2. �ɰ����������е���˿������˿�������λ��CRTs��ģʽ��A-C��Ⱦɫ������ģʽ�ĵ��Ͱ�����������֮��ɼ����Է�ͬԴȾɫ������λ���Ҷϵ�����������˿����ͼ�л�ɫ���ߣ���Hi-C�ź�ȷ�ϣ�������A�����ɼ���������ˮɼ���ؿ�����ȣ�����Ⱦɫ�嶼������������ϣ��Ҷϵ�����������˿��������B�����ɼ���������������ȣ�7��Ⱦɫ�巢����λ���ϵ����ɵ���˿����������ƫ�������ӵ���˿����C����D���о��Ŷ�����������CRTsģ������������������Ⱦɫ������ģʽ������ͼ�۵ķ�ʽ������·����E-F��A-C��Ӧ��Ⱦɫ�������ͼ��

ͼ3. �ؽ�������ֲ������ݻ���ʷ��A���Ʋ������ֲ����Ҫ��֧����������������ĺ����ݻ���ʷ��B����Ŀ��һ������˿��Ⱦɫ���ڲ�ͬ��֧�е��ݻ���ʷ���Լ��챶��(dysploidy)����;����
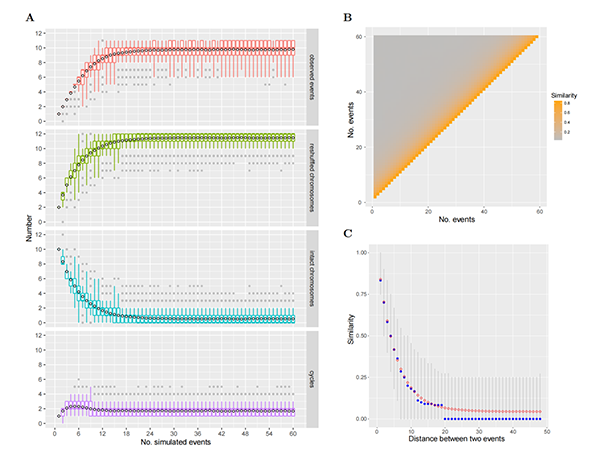
ͼ4. CRTģ�͵�ģ�⡣������Ⱦɫ����Ŀ��n?= 12���£���CRT������������ʱ���ɹ۲쵽��CRT�¼����ڱ��͡�
����ͨ�Ź��ں�
֪����ҵ��Ƹ

