
-

生物通官微
陪你抓住生命科技
跳动的脉搏
生物通官微
陪你抓住生命科技
跳动的脉搏
华南植物园发现系统发育基因组学和形态学证据支持亚洲产崖豆藤属划分为3个属
【字体: 大 中 小 】 时间:2024年12月17日 来源:中国科学院华南植物园
编辑推荐:
中国科学院华南植物园助理研究员宋柱秋与合作者,从2013年开始着手研究,基于大量的馆藏标本、野外调查和分子系统学研究,陆续获得该属在分类上的一些进展,主要包括:(1)建立一个新属——启明豆属Huchimingia(Song et al. 2022a. Phytotaxa 532: 37-56),仅5种,包括此前描述的1个乔木新种韦直启明豆(Song et al. 2019. Nordic Journal of Botany 35: 404-410),以及1个中国新记录种柄果启明豆;(2)将部分种转移或归并至至鱼藤属Derris,如厚果崖豆藤、榼藤子崖豆藤、Millettia boniana等?(Song & Pan 2022. Phytotaxa 531: 230-248);(3)恢复白崖豆属Imbraylx的属级地位(Song 2021. Phytotaxa 525: 223-231),该属隶属于核心崖豆藤族的刀豆氨酸群(Canavanine group),区别于崖豆藤属所在的非刀豆氨酸群(Non-canavanine group);(4)将无患子崖豆藤归并至云南鱼藤,并首次对此种的系统位置提出质疑(Song et al. 2017a. Nordic Journal of Botany 35: 404-410),后来该种被其他研究者抢先发表为新属——异齿豆属Ohashia;(5)证明澜沧崖豆藤是一个混杂的种,花标本属于草崖豆藤Cruddasia insignis,果标本属于球子鸡血藤Callerya sphaerosperma,由于花标本被指定为模式标本,因此将该种归并到草崖豆藤,由此报道草崖豆藤属为中国新记录属(Song et al. 2021a. Phytotaxa 497: 29-38);(6)综述了越南和印度产崖豆藤属的现状(Song et al. 2022b. Phytotaxa, 571, 169-185; Song 2023a. Phytotaxa 591: 55-63),澄清了该属其他一些种级水平的分类问题(Song et al. 2017b. Phytotaxa 32: 51-58; Song et al. 2021b. Phytotaxa 507: 183-190);(7)也对崖豆藤属的近缘属——短翅鱼藤属进行了分类学研究(Song 2023b. Phytotaxa 484: 251-263)
崖豆藤属Millettia隶属于豆科蝶形花亚科核心崖豆藤族(core Millettieae)。该属是一个分类复杂的大属,全世界约有150种,分布于亚洲和非洲。以往的研究已经表明崖豆藤属在形态上高度多样,分子证据也证明该属是高度多系。2005年,英国邱园出版的《世界豆类》被誉为豆科植物的百科全书,该专著明确指出崖豆藤属的分类迫切需要大修(major revision)。
中国科学院华南植物园助理研究员宋柱秋与合作者,从2013年开始着手研究,基于大量的馆藏标本、野外调查和分子系统学研究,陆续获得该属在分类上的一些进展,主要包括:(1)建立一个新属——启明豆属Huchimingia(Song et al. 2022a. Phytotaxa 532: 37-56),仅5种,包括此前描述的1个乔木新种韦直启明豆(Song et al. 2019. Nordic Journal of Botany 35: 404-410),以及1个中国新记录种柄果启明豆;(2)将部分种转移或归并至至鱼藤属Derris,如厚果崖豆藤、榼藤子崖豆藤、Millettia boniana等?(Song & Pan 2022. Phytotaxa 531: 230-248);(3)恢复白崖豆属Imbraylx的属级地位(Song 2021. Phytotaxa 525: 223-231),该属隶属于核心崖豆藤族的刀豆氨酸群(Canavanine group),区别于崖豆藤属所在的非刀豆氨酸群(Non-canavanine group);(4)将无患子崖豆藤归并至云南鱼藤,并首次对此种的系统位置提出质疑(Song et al. 2017a. Nordic Journal of Botany 35: 404-410),后来该种被其他研究者抢先发表为新属——异齿豆属Ohashia;(5)证明澜沧崖豆藤是一个混杂的种,花标本属于草崖豆藤Cruddasia insignis,果标本属于球子鸡血藤Callerya sphaerosperma,由于花标本被指定为模式标本,因此将该种归并到草崖豆藤,由此报道草崖豆藤属为中国新记录属(Song et al. 2021a. Phytotaxa 497: 29-38);(6)综述了越南和印度产崖豆藤属的现状(Song et al. 2022b. Phytotaxa, 571, 169-185; Song 2023a. Phytotaxa 591: 55-63),澄清了该属其他一些种级水平的分类问题(Song et al. 2017b. Phytotaxa 32: 51-58; Song et al. 2021b. Phytotaxa 507: 183-190);(7)也对崖豆藤属的近缘属——短翅鱼藤属进行了分类学研究(Song 2023b. Phytotaxa 484: 251-263)。但是已有的这些研究依然未能解决亚洲产崖豆藤属多数种的归属问题,该属剩余的亚洲三个组也均不是单系类群,需要进一步研究。
近期,基于第二代测序数据提取了完整的叶绿体基因组和2000余个单拷贝核基因,在核心崖豆藤族完整属级取样的情况下,解决了亚洲产崖豆藤属的分类。该研究主要结果包括:(1)重新界定了核心崖豆藤族的范围,正式将菜豆族拟大豆亚族的3个属(Cruddasia,?Ophrestia和Pseudoeriosema)纳入核心崖豆藤族的范围,将核心崖豆藤族分为2群(刀豆氨酸群和非刀豆氨酸群)5个分支及14个亚支,总共包含29个属约1000种;(2)亚洲产崖豆藤属种类聚成3个高支持率的亚支,并得到形态证据的支持,因此这三个亚支被处理为三个属,即水黄皮属Pongamia(56种)、耳旗豆属Otosema(3种)和狭义崖豆藤属Millettia?s.str.(7种),并将干花豆属Fordia和糠皮豆属Ibatiria归并到水黄皮属,共建立了56个新组合和2个新名称。(3)基于分化时间估计,推测核心崖豆藤族的一个主要分支(clade VII)经历了早期的快速分化,该分支涉及崖豆藤族的三个长期有分类争议的中心属,即广义崖豆藤属、广义鱼藤属Derris?s.lato和广义醉鱼豆属Lonchocarpus?s.lato。因此推测这种基于形态上的有争议的分类处理,可能与快速分化导致的复杂的形态性状演化有关。
相关研究成果已近期发表在国际知名期刊Molecular Phylogenies and Evolution(《分子系统发育与进化》)上。华南植物园宋柱秋为第一作者,合作者包括华南植物园李世晋研究员,徐连升助理研究员,科研助理蒋凯文,复旦大学赵义勇博士,武汉植物园李湉博士,印度马拉巴尔植物园和植物科学研究所A.P. Balan博士,以及法国国家自然历史博物馆M. Gaudeul博士。该研究得到了广东省基础与应用基础研究旗舰项目的支持。论文链接:https://www.sciencedirect.com/science/article/abs/pii/S105579032400246X
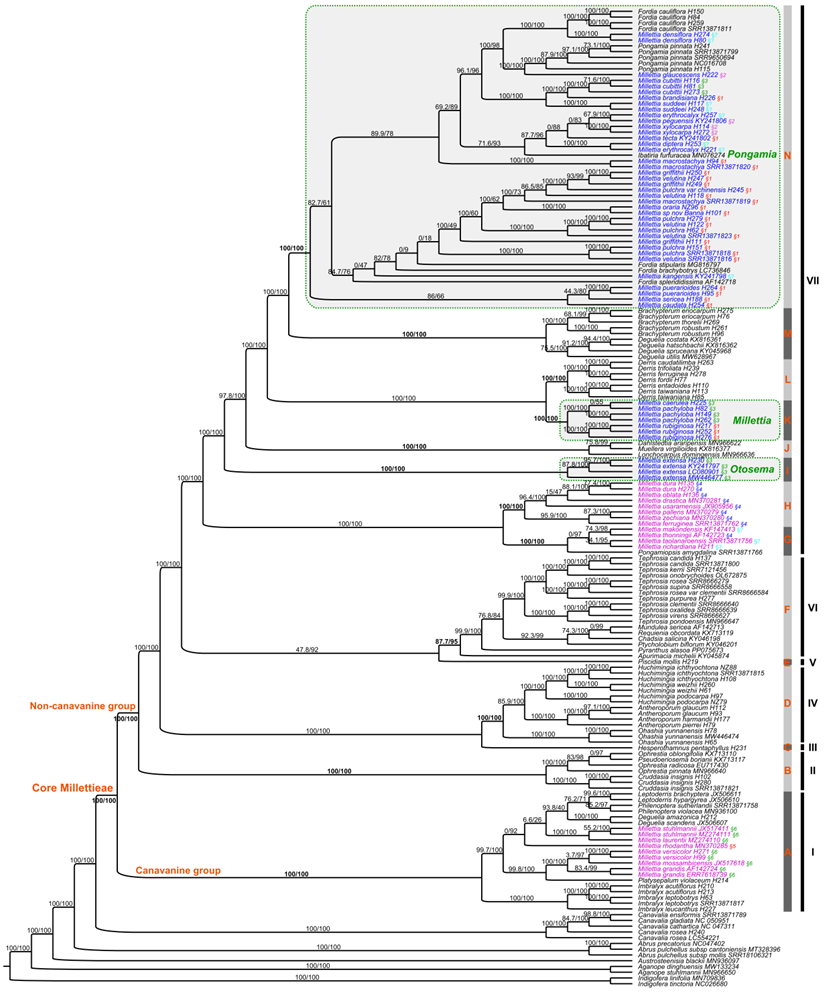
图1. 基于完整叶绿体基因组的核心崖豆藤族的系统发育关系
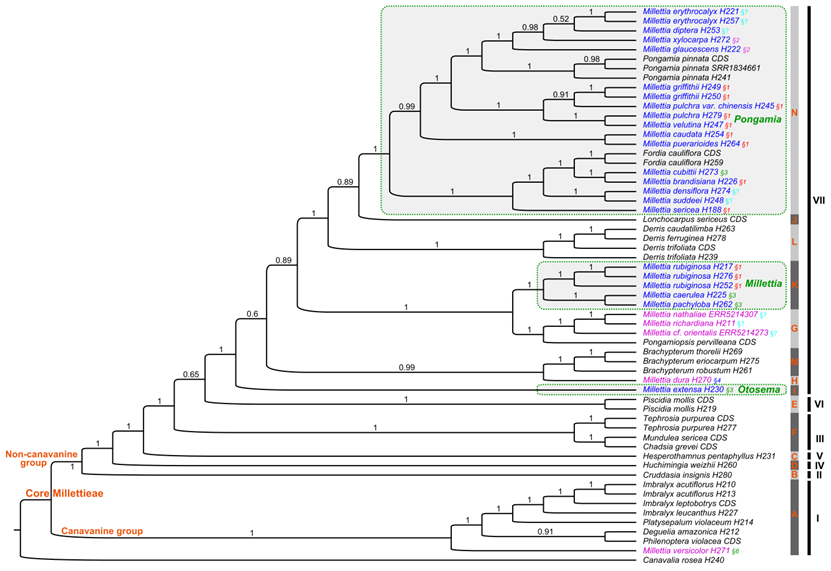
图2. 基于2106个单拷贝核基因的核心崖豆藤族的系统发育关系
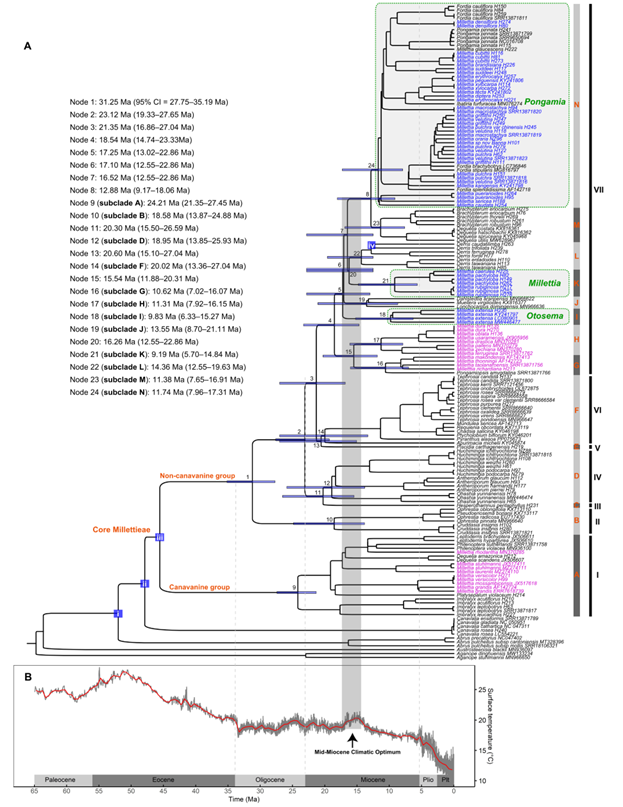
图3. 核心崖豆藤族的分化时间估计

