
-

生物通官微
陪你抓住生命科技
跳动的脉搏
生物通官微
陪你抓住生命科技
跳动的脉搏
朱学良组合作揭示上皮组织运动纤毛协调摆动的调控机制
【字体: 大 中 小 】 时间:2024年11月28日 来源:中国科学院生物化学与细胞生物学研究所
编辑推荐:
11月26日,国际学术期刊Nature?Communications在线发表了中国科学院分子细胞科学卓越创新中心(生物化学与细胞生物学研究所)朱学良研究组联合上海交通大学医学院附属新华医院鄢秀敏课题组的最新合作研究成果“Directional?ciliary beats across epithelia require Ccdc57-mediated coupling between axonemal?orientation and basal body polarity”
11月26日,国际学术期刊Nature?Communications在线发表了中国科学院分子细胞科学卓越创新中心(生物化学与细胞生物学研究所)朱学良研究组联合上海交通大学医学院附属新华医院鄢秀敏课题组的最新合作研究成果“Directional?ciliary beats across epithelia require Ccdc57-mediated coupling between axonemal?orientation and basal body polarity”。该研究揭示了上皮组织运动纤毛协调摆动的新调控机制。
运动纤毛是一种进化上高度保守的毛状细胞器,其核心骨架轴丝(axoneme)是由九组二联体微管和一对中央微管组成的、具有明确朝向(axonemal?orientation, AO)的旋转不对称结构。轴丝的二联体微管与锚定在细胞膜上的基体(basal?body)相连,而轴丝上附着的包括分子马达在内的各种蛋白质复合物使纤毛能沿轴丝方向进行周期性的来回摆动。在哺乳动物中,运动纤毛以多纤毛(每个细胞数十到数百根)的方式广泛分布于脑室、气管和输卵管等上皮组织表面。这些纤毛初始的摆动方向是混乱的,它们需要在发育中逐渐建立平面细胞极性(planar?cell polarity,?PCP),即形成组织范围内协调统一的摆动方向。例如,小鼠脑室管膜上皮细胞的多纤毛在小鼠出生3天左右才发生,到21天龄以后才能建立PCP。这样缓慢的进程提示其中过程的复杂性。协调摆动的纤毛驱使脑脊液、痰液等组织外液定向流动,以促进中枢神经系统稳态维持、呼吸道清洁和受精卵向子宫的运动。运动纤毛结构和功能的缺陷,会导致主要病征包括慢性呼吸系统疾病、不孕不育、内脏倒位和脑积水的原发性纤毛运动障碍(primary?ciliary dyskinesia)。早期研究发现,纤毛的摆动方向还与基体的极性,即基体附属结构基足(basal foot,?BF)的指向相偶联。因此,通常认为在纤毛建立PCP的过程中,纤毛摆动方向的改变是细胞骨架通过牵拉BF的顶端而旋转整根纤毛的结果。然而,AO如何与基体极性相偶联却是领域中的空白。而且,如果偶联不是从纤毛形成之初就发生,那现有的理论认知或许就需要予以修正。
该研究团队发现Ccdc57的进化保守性与上皮组织运动纤毛密切相关,提示它可能是纤毛PCP的重要调节因子。有趣的是,在小鼠脑室管膜上皮细胞中,Ccdc57呈点状以旋转不对称的方式定位在基体远端的管腔内,而且伴随着纤毛PCP的建立,该定位从随机变为极化到BF对侧。Ccdc57基因敲除小鼠出现严重的脑积水和高死亡率。深入的分析发现,敲除小鼠脑室管膜上皮细胞内不同纤毛能形成协调一致的摆动,但不同细胞间却缺乏统一,这导致脑脊液流动异常而引起脑积水和死亡。而且,Ccdc57敲除纤毛的AO与BF是解偶联的,因此摆动方向与基体极性缺乏相关性。随后,研究人员分析了4天龄野生型小鼠细胞中摆动方向混乱的纤毛,发现其多数的AO与BF是未偶联的,而在摆动方向一致的纤毛中AO-BF偶联的则占多数。研究人员还分析了Ccdc57敲除小鼠气管上皮的多纤毛,发现其AO-BF偶联也存在缺陷。这些研究结果揭示AO与BF并非从动纤毛一发生就已偶联,并说明细胞内的纤毛簇可以不依赖于其基体极性的方式形成一致性的摆动,但需要偶联形成后才能建立PCP。而且,研究人员发现了首个介导AO-BF偶联的关键因子Ccdc57,并提示在基体远端存在一个包含该蛋白质的新结构,负责在纤毛建立PCP的过程中锁定轴丝和基体的极性关系。这些发现修订了领域内先前的认知,并为理解运动纤毛PCP的调节机理和相关病理机制提供了新视角。
分子细胞卓越中心与上海科技大学联合培养博士生潘新文、分子细胞卓越中心博士生方楚玉、分子细胞卓越中心与上海科技大学联合培养硕士生沈川为共同第一作者。分子细胞卓越中心朱学良研究员和上海交通大学医学院附属新华医院鄢秀敏研究员为该论文的共同通讯作者。中国科学院生物物理研究所李喜霞高级工程师等研究人员为合作者。该研究获得国家自然科学基金委、科技部和中国科学院的资助,以及中国科学院生物物理研究所生物成像中心,分子细胞卓越中心细胞分析技术平台、分子生物学技术平台,国家蛋白质设施等的技术支持。
文章链接:https://www.nature.com/articles/s41467-024-54766-1
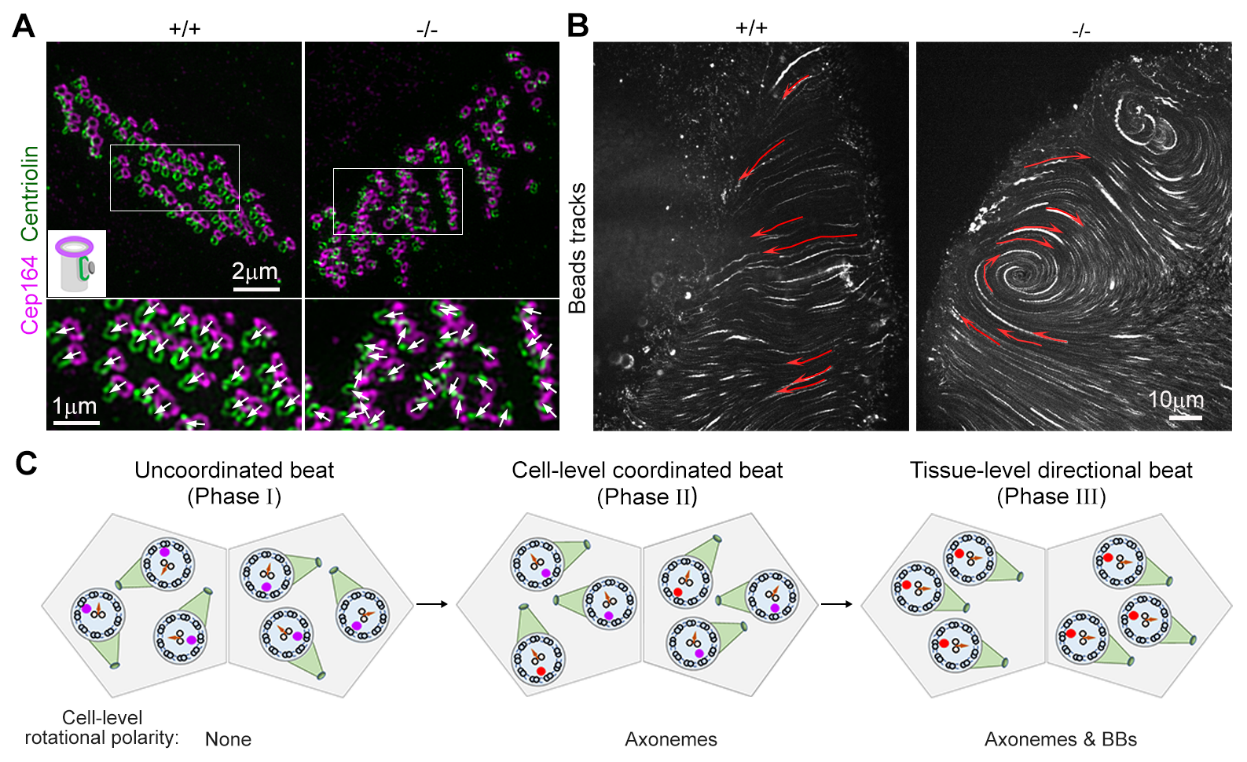
(A)相对野生型(+/+)小鼠,Ccdc57敲除(-/-)小鼠脑室管膜细胞多纤毛的基体不能形成一致的极性。Cep164标记基体,而Centriolin标记基足,箭头表示基足方向。(B)Ccdc57敲除导致脑室管膜上皮细胞纤毛驱动的液体流动缺乏定向性(红色箭头)。(C)模型图展示新生纤毛的轴丝方向(箭头)与基足(绿色)在阶段I未偶联,此时Ccdc57呈现随机的点状定位(紫红)。此时细胞内多纤毛的摆动方向是混乱的。在阶段II中,细胞内的轴丝方向逐渐统一,其中一部分与基足偶联,Ccdc57也相应呈现极性定位(红色)。这时同一细胞内的多纤毛实现定向摆动,但不同细胞缺乏一致性。在阶段III中,轴丝方向与基体极性完成偶联,从而实现纤毛在组织层面上的定向摆动(PCP)。

