
-

…˙ŒÔÕ®πŸŒ¢
≈„ƒ„◊•◊°…˙√¸ø∆ºº
ï∂صƒ¬ˆ≤´
…˙ŒÔÕ®πŸŒ¢
≈„ƒ„◊•◊°…˙√¸ø∆ºº
ï∂صƒ¬ˆ≤´
…˙√¸ø∆—ß—ß‘∫÷İ—ŞøŒÃ‚◊È‘ĞCell Research…œ¡™∫œ∑¢ŒƒΩ“ æ≤°‘≠æ˙±Ş√´¬Ì¥Ô–˝◊™∑ΩœÚ◊™ªªµƒ∑÷◊”ª˙÷∆
°æ◊÷ã∫ ¥Û ÷– –° °ø ±º‰£∫2024ƒÍ10‘¬13»’ ¿¥‘¥£∫’„Ω≠¥Û—ß…˙√¸ø∆—ß—ß‘∫
±‡º≠Õ∆ºˆ£∫
°°°°2024ƒÍ8‘¬23»’£¨Œ“‘∫÷İ—ŞøŒÃ‚◊È∫Õ…˙—–‘∫÷Ï”¿»∫øŒÃ‚◊È¡™∫œ‘ĞCell Research‘”÷æ‘Ğœş∑¢±Ì¡ÀÂŒ™ °∞Structural basis of the bacterial flagellar motor rotational switching°±µƒ—–æø¬ğŒƒ£¨Õ®π˝ππΩ®º§ªÓ–Õ«˜ªØ“Ú◊”µ∞∞◊CheYÕª±‰Ã£¨¥øªØ¿¥‘¥”Ğ≤°‘≠æ˙…≥√≈ œæ˙£®Salmonella Typhimurium£©µƒƒĞ‘¥–‘µƒ°¢∫¨∑ΩœÚø™πÿ∏¥∫œŒÔ∞˚÷ ª∑£®C ring£©µƒ±Ş√´¬Ì¥Ôø≈¡££¨Ω‚Œˆ¡À∑÷±¥¶”ĞƒÊ ±’Î∫ÕÀ≥ ±’Ζ˝◊™∑ΩœÚ÷–µƒ°¢ÕÍ’˚µƒ±Ş√´¬Ì¥Ô-Ω”Õ∑◊∞÷√∏¥∫œŒÔµƒ...
2024ƒÍ8‘¬23»’£¨Œ“‘∫÷İ—ŞøŒÃ‚◊È∫Õ…˙—–‘∫÷Ï”¿»∫øŒÃ‚◊È¡™∫œ‘ĞCell Research‘”÷æ‘Ğœş∑¢±Ì¡ÀÂŒ™ °∞Structural basis of the bacterial flagellar motor rotational switching°±µƒ—–æø¬ğŒƒ£¨Õ®π˝ππΩ®º§ªÓ–Õ«˜ªØ“Ú◊”µ∞∞◊CheYÕª±‰Ã£¨¥øªØ¿¥‘¥”Ğ≤°‘≠æ˙…≥√≈ œæ˙£®Salmonella Typhimurium£©µƒƒĞ‘¥–‘µƒ°¢∫¨∑ΩœÚø™πÿ∏¥∫œŒÔ∞˚÷ ª∑£®C ring£©µƒ±Ş√´¬Ì¥Ôø≈¡££¨Ω‚Œˆ¡À∑÷±¥¶”ĞƒÊ ±’Î∫ÕÀ≥ ±’Ζ˝◊™∑ΩœÚ÷–µƒ°¢ÕÍ’˚µƒ±Ş√´¬Ì¥Ô-Ω”Õ∑◊∞÷√∏¥∫œŒÔµƒ¡Ω∏ˆ∏ş∑÷±Ê¬ ¿‰∂≥µÁæµΩ·ππ£¨«ÂŒ˙µÿΩ“ æ¡ÀCheYµ∞∞◊Ω·∫œµº÷¬∑ΩœÚø™πÿ∏¥∫œŒÔµƒππœÛ±‰ªØ£¨Ã·≥ˆ¡À±Ş√´¬Ì¥Ô∂®◊”µ•‘™µƒƒĞƒ§÷ÿ∂®Œª∏≈ƒÓ£¨¥”∂¯≤˚√˜¡Àœ∏æ˙±Ş√´¬Ì¥Ô–˝◊™∑ΩœÚ◊™ªªµƒ∑÷◊”ª˙÷∆£¨æ¿’˝¡À÷Æ«∞À˘”–πÿ”ĞC ringµƒ◊È◊∞∫ÕΩ·ππµƒ¥ÌŒÛ¿ÌΩ‚£¨»À√«’π æ¡ÀÕÍ’˚µƒœ∏æ˙±Ş√´¬Ì¥ÔµƒΩ·ππº∞π§◊˜‘≠¿Ì°£
œ∏æ˙±Ş√´ª˘±æΩ·ππ∞¸¿®±Ş√´Àø£®filament£©°¢Ω”Õ∑◊∞÷√£®hook£©“‘º∞±Ş√´¬Ì¥Ô£®flagellar motor£©»˝≤ø∑÷°£±Ş√´¬Ì¥Ô «”…œ‚«∂‘Ğœ∏æ˙œ∏∞˚ƒ§…œµƒ◊™◊”£®rotor£©º∞∆‰÷İŒß√™∂®µƒ∂®◊”µ•‘™£®stator units£©◊È∫œ∂¯≥…°£∂®◊”µ•‘™ «“ª÷÷¿Î◊”Õ®µ¿£¨ø…“‘¿˚”√œ∏æ˙œ∏∞˚ƒ§ƒĞÕ‚µƒ¿Î◊”µÁªØ—ßÃı∂»º¥¿Î◊”∂Ø¡¶ ∆¥Ÿ π∂®◊”µ•‘™∑¢…˙–˝◊™£¨Ω´ªØ—߃İ◊™ªØŒ™ª˙–µƒİ¿¥≤˙…˙≈§æÿ£¨≈§æÿæ≠◊™◊”Ω·ππ÷–Œª”Ğ∞˚÷ “ª≤‡µƒ∞˚÷ ª∑£®cytoplasmic ring£¨C ring£©¥´µıµΩ¡™∂Ø∏À£¨‘ŸΩ¯∂¯¥´µı∏¯Ω”Õ∑◊∞÷√∫Õ‘∂∂À±Ş√´Àø£¨¥”∂¯«˝∂Øœ∏æ˙‘À∂Ø°£
œ∏æ˙±Ş√´¬Ì¥Ô «“ª∏ˆæŞ¥ÛµƒƒİÀ´œÚ–˝◊™µƒ∑÷◊”¬Ì¥Ô£¨Õ®∏≥”Ë¡Àœ∏æ˙≥¨º∂‘À∂؃ݡ¶£¨ƒİ«˝ πœ∏æ˙√ø√Î÷””Œ∂Ø≥§¥Ô◊‘º∫…Ì≥§º∏ Ʊ∂°¢…÷¡…œ∞Ÿ±∂µƒæ‡¿Î°£≤ªΩˆ»Á¥À£¨œ∏æ˙±Ş√´¬Ì¥Ôƒİ‘ĞƒÊ ±’Î∫ÕÀ≥ ±’Ζ˝◊™∑ΩœÚ÷ƺ‰Ω¯––◊™ªª°£’˝≥£«Èøˆœ¬±Ş√´ƒÊ ±’Ζ˝◊™£¨œ∏æ˙œÚ«∞”Œ∂Ø°£µ±«˜ªØ–≈∫≈◊™µºÕ®¬∑÷–CheYµ∞∞◊ª·±ª¡◊À·ªØ£¨¡◊À·ªØµƒCheY£®CheY-P£©Õ®π˝Ω·∫œµΩC ring£¨¥Ÿ π±Ş√´¬Ì¥Ô–˝◊™∑ΩœÚ¥”ƒÊ ±’Σ®CCW£©◊™ªªŒ™À≥ ±’Σ®CW£©∑ΩœÚ£¨◊Ó÷’∏ƒ±‰æ˙ÑÀ∂Ø∑ΩœÚ£¨“Ú¥ÀC ring“≤±ª≥∆Œ™∑ΩœÚø™πÿ∏¥∫œŒÔ£®switch complex£©°£¥ÀÕ‚£¨CheZø…“‘Ω´CheY-P»•¡◊À·ªØΩ¯∂¯∑¥µ˜øÿ¬Ì¥Ôµƒ–˝◊™∑ΩœÚ°£”…”ĞC ringΩ·ππ∏ş∂»∂Øè«“‘ЃБ¥–‘¥øªØ ±»ı“◊¥”±Ş√´¬Ì¥Ô…œΩ‚æ𣨓ڥÀæğΩπ”ĞC ringµƒ±Ş√´¬Ì¥Ô–˝◊™∑ΩœÚµ˜øÿµƒΩ·ππª˙÷∆—–æø“ª÷±¥Ê‘Ğ≤ª…Ÿµƒ’œ∞≠£¨µº÷¬œ∏æ˙±Ş√´¬Ì¥Ô–˝◊™∑ΩœÚ◊™ªªµƒ∑÷◊”ª˙÷∆≥§∆Ğ≤ª«Â≥˛£¨ «¡Ï”ÚƒĞ≥§∆Ğ–¸∂¯Œ¥æˆµƒ÷ÿ“™Œ Â÷Æ“ª°£

øŒÃ‚Õ≈∂”◊‘2021ƒÍ‘ĞCell‘”÷æ∑¢±Ì¬ğŒƒΩ“ æœ∏æ˙±Ş√´¬Ì¥Ô◊È◊∞∫Õ≈§æÿ¥´ ‰ª˙÷∆“‘¿¥£¨“ª÷±—–æøœ∏æ˙±Ş√´¬Ì¥Ôπ§◊˜ª˙÷∆£¨≤ª∂œ”≈ªØ…≥√≈ œæ˙ƒĞ‘¥–‘±Ş√´¬Ì¥Ô¥øªØÃÂœµ£¨Õ®π˝ππΩ®º§ªÓ–ÕCheYÕª±‰Ã£®CheY**£¨º¥CheYD13K&Y106WÕª±‰Ã£©£¨≥…π¶ªÒµ√¡ÀƒÊ ±’Î∫ÕÀ≥ ±’Ζ˝◊™◊¥Ã¨œ¬…≥√≈ œæ˙∫¨C ringµƒÕÍ’˚±Ş√´¬Ì¥Ô-Ω”Õ∑◊∞÷√∏¥∫œŒÔø≈¡££¨≤¢Ω‚Œˆ¡ÀÀ¸√«µƒ¿‰∂≥µÁæµΩ·ππ£®Õº1£©°£∫¨CCW-C ringµƒ±Ş√´¬Ì¥Ô-Ω”Õ∑◊∞÷√∏¥∫œŒÔΩ·πππ≤∞¸∫¨341∏ˆ—«ª˘£¨”…15÷÷≤ªÕ¨±Ş√´µ∞∞◊◊È≥…£¨’˚ÃÂ∑÷◊”¡ø‘ºŒ™10.2MDa£¨∏ş∂»‘ºŒ™680?°£∂¯∫¨CheY**Ω·∫œµƒCW-C ringµƒ±Ş√´¬Ì¥Ô-Ω”Õ∑◊∞÷√∏¥∫œŒÔ’˚∏ˆƒ£–Õ”…375∏ˆ—«ª˘◊È≥…£¨∏ş∂»‘ºŒ™664?°£¡Ω÷÷–˝◊™◊¥Ã¨œ¬µƒC ringæ˘æş”–C34∂‘≥∆–‘£¨”…34∏ˆFliG°¢34∏ˆFliM∫Õ102∏ˆFliN“‘1:1:3µƒƒ¶∂˚±»¿˝◊È≥…£¨ƒĞ≤øæş”–π„∑∫«“∂¿Ãÿµƒ—«ª˘º‰µƒΩª≤ʪ•◊˜£¨ø…∑÷Œ™ƒĞ≤ø—«ª∑°¢…œ≤ø—«ª∑°¢÷–º‰—«ª∑∫Õµ◊≤ø—«ª∑π≤º∆Àƒ∏ˆ—«ª∑°£∆‰÷–£¨FliGÕ®π˝∂—ª˝÷¬∆‰C∂ÀΩ·ππ”ÚFliGCC≈≈¡––Œ≥…¡Àø…“‘”Î∂®◊”µ•‘™ª•◊˜µƒ…œ≤ø—«ª∑Ω·ππ°£CheY**µƒΩ·∫œµº÷¬¡ÀC ringµƒ’˚ÃÂΩ·ππ∑¢…˙¡À√˜œ‘µƒ«„–±∫՜ڃР’À£¨≤¢œÚMS ring∫Õœ∏æ˙ƒĞƒ§øøΩ¸16 ?µƒæ‡¿Î£®Õº1£©°£
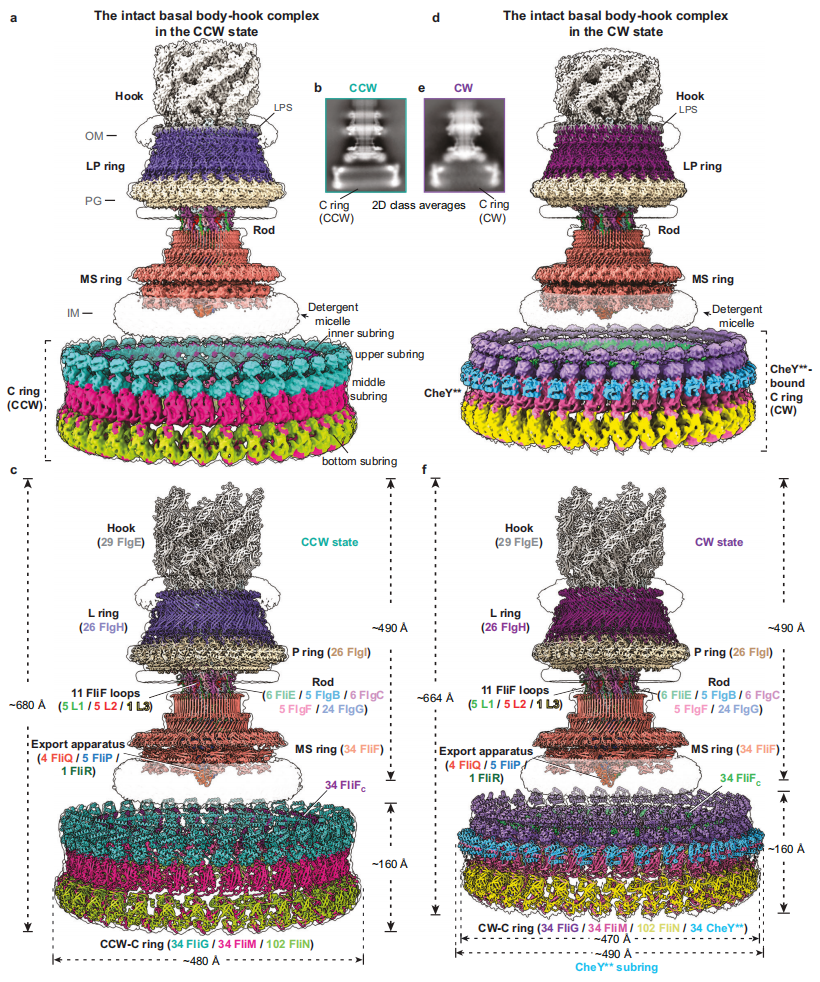
Õº1. ∫¨∑ΩœÚø™πÿ∏¥∫œŒÔµƒœ∏æ˙±Ş√´¬Ì¥Ô‘ĞƒÊ ±’Î∫ÕÀ≥ ±’Ζ˝◊™◊¥Ã¨œ¬µƒ¿‰∂≥µÁæµµÁ◊”√İ∂»∫Õ»˝Œ¨Ω·ππ
º§ªÓ–ÕCheYÕ®π˝FliMµƒN∂ÀΩ·ππ”ÚΩ·∫œ‘Ğœ‡¡ĞFliMMº∞FliGMΩ·ππ”ÚÕ‚±Ì√Ê–Œ≥…µƒº‰œ∂÷–£¨µº÷¬C ringµƒ∏˜Ω·π𵕑™æ˘≥ˆœ÷≤ªÕ¨≥Ã∂»µƒππœÛ±‰ªØ£¨Ãÿ± «FliG—«ª˘µƒππœÛ∑¢…˙¡ÀæŞ¥Û±‰ªØ£®Õº2£©°£CheYµƒΩ·∫œ∏ƒ±‰¡ÀFliG—«ª˘À˘”–Ω·ππ”Úµƒø’º‰Œª÷√∫ÕΩ·ππ”Úº‰µƒœ‡∂‘懿Σ¨Ω¯∂¯µº÷¬FliGCCΩ·ππ”Ú∑¢…˙¡À180°„–˝◊™≤¢œÚ¬Ì¥Ô÷––ƒ“∆∂Ø¡À10 ?µƒæ‡¿Î£¨¥Ÿ π”Î∂®◊”µ•‘™ª•◊˜µƒFliGCCΩ·ππ”Úµƒ∫À–ƒ¬ı–˝Ω·ππ¶¡torque≥ˆœ÷»°œÚ∫Õæ≤µÁ ∆ƒİ∑÷≤ºµƒ∑≠◊™°£C ring µƒππœÛ±‰ªØÀµ√˜∂®◊”µ•‘™–Μڬ̥Ô÷––ƒ“∆∂Ø£¨–Ë“™‘Ğœ∏æ˙ƒĞƒ§…œ∑¢…˙÷ÿ–¬∂®Œª£¨“‘ ”¶¶¡torqueø’º‰Œª÷√µƒ±‰ªØ°£Õ®π˝∑÷Œˆ“‘Õ˘±®µ¿π˝µƒ¿‰∂≥∂œ≤„…®√Ë ˝æı£¨Ω·∫œΩ®¡¢∫¨∂®◊”µ•‘™µƒÕÍ’˚±Ş√´¬Ì¥ÔΩ·ππƒ£–Õ£®Õº3£©£¨÷§√˜¡À∂®◊”µ•‘™»∑ µ‘Ğœ∏æ˙ƒĞƒ§…œ∑¢…˙÷ÿ–¬∂®Œª°£

Õº2. º§ªÓ–ÕCheYΩ·∫œµº÷¬C ring∑¢…˙ππœÛ±‰ªØ
“‘…œ ˝æı∑÷Œˆ’π æ¡À±Ş√´¬Ì¥Ô∑ΩœÚ◊™ªªµƒ∑÷◊”ª˙÷∆£∫‘ĞŒŞCheYµ∞∞◊Ω·∫œ ±£¨—ÿÀ≥ ±’Î∑ΩœÚ–˝◊™µƒ∂®◊”µ•‘™Œª”ĞC ringµƒFliGCC—«ª∑µƒÕ‚≤‡£¨«˝∂رŞ√´¬Ì¥Ô—ÿƒÊ ±’Î∑ΩœÚ–˝◊™°£µ±CheYΩ·∫œµΩC ring ±£¨¥ŸΩ¯C ring ∑¢…˙ππœÛ±‰ªØ£¨œÚ±Ş√´¬Ì¥Ô÷––ƒ ’À≤¢’˚ÃÂ…œ“∆£¨Ω¯∂¯“˝∆FliGCCΩ·ππ”ÚππœÛ±‰ªØ£¨◊Ó÷’µº÷¬FliGCC…œµƒ¶¡torque∑¢…˙¡À180°„µƒ∑≠◊™£¨Ω¯∂¯ µœ÷∂®◊”µ•‘™¥”FliGÕ‚≤‡◊™µΩƒĞ≤‡£¨‘ЃЃ§…œ∑¢…˙ø’º‰÷ÿ∂®Œª£¨¥Ÿ πC ring“‘À≥ ±’Î∑ΩœÚΩ¯–––˝◊™£¨¥”∂¯ µœ÷±Ş√´¬Ì¥Ô∑ΩœÚµƒ◊™ªª£®Õº3£©°£
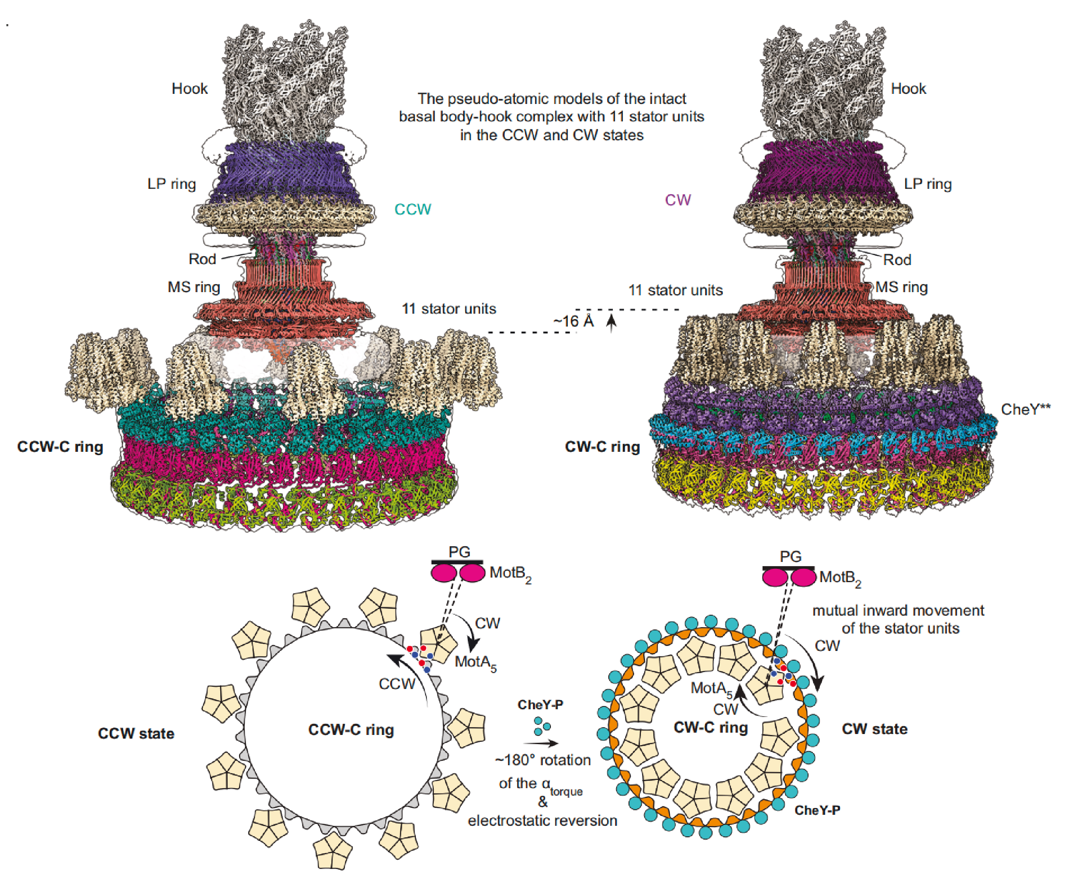
Õº3. œ∏æ˙±Ş√´¬Ì¥Ô∑ΩœÚ◊™ªªµƒª˙÷∆ƒ£ ΩÕº
’‚œÓ—–æøπ§◊˜æ¿’˝¡À÷Æ«∞À˘”–πÿ”ĞC ringµƒ◊È◊∞∫ÕΩ·ππµƒ¥ÌŒÛ¿ÌΩ‚£¨∏ƒ±‰¡À÷Æ«∞»œŒ™∂®◊” «ÕÍ»´πÃ∂®‘Ğœ∏æ˙ƒĞƒ§…œµƒ∏≈ƒÓ£¨∑¥≤µ¡À“‘Õ˘À˘”–µƒπÿ”бŞ√´¬Ì¥Ô∑ΩœÚ◊™ªª «“ÚŒ™C ringµƒΩ·ππÕ‚—”’‚“ªºŸÀµ£¨≤¢œÚ»À√«’π æ¡ÀÕÍ’˚±Ş√´¬Ì¥Ô»˝Œ¨Ω·ππ∫Õπ§◊˜ª˙÷∆£¨Œ™…˺∆–¬øπæ˙“©ŒÔ“‘º∞–¬–Õ–˝◊™ƒ…√◊ª˙∆˜»ÀµÏ∂®¡À¿Ì¬ğª˘¥°°£
÷İ—Ş—–æø‘±∫Õ÷Ï”¿»∫Ωà Р«±æŒƒµƒπ≤Õ¨Õ®—∂◊˜’ş£¨Ã∑º”–À≤© ø∫Õ’≈¡·≤© øŒ™±æŒƒµƒπ≤Õ¨µĞ“ª◊˜’ş£¨≤Œº”—–æøµƒªπ”–—–æø…˙÷İ–«ÕÆ∫Õ∫´À∆”Òµ»°£’„Ω≠¥Û—ß°¢∏¥µ©¥Û—ß°¢…œ∫£ΩªÕ®¥Û—ß“Ω—ß‘∫æ´◊º“Ω—ß—–æøÀ˘µ»µ•Œªµƒ¿‰∂≥µÁæµ÷––ƒ≤Œ”εÁæµ ˝æıµƒ ’ºØπ§◊˜°£±æ—–æø”…π˙º“◊‘»ªø∆—ߪ˘Ω°¢π˙º“÷ÿµ„—–∑¢º∆ªÆ∫Õ÷–—Î∏ş–£ª˘±æø∆—–“µŒÒ∑—µƒ◊ ÷˙°£
‘≠Œƒ¡¥Ω”£∫https://www.nature.com/articles/s41422-024-01017-z
…˙ŒÔÕ®Œ¢–≈π´÷Ğ∫≈
÷™√˚∆Û“µ’–∆∏
ΩÒ»’∂Øè | »À≤≈ –≥° | –¬ºº ◊®¿∏ | ÷–π˙ø∆—ß»À | ‘∆’πî | BioHot | ‘∆Ω≤Ã√÷±≤• | ª·’π÷––ƒ | Ãÿºğ◊®¿∏ | ºº øÏ—∂ | √‚∑— ‘”√
∞Ê»®À˘”– …˙ŒÔÕ®
Copyright© eBiotrade.com, All Rights Reserved
¡™œµ–≈œ‰£∫
‘¡ICP±∏09063491∫≈
