
-

生物通官微
陪你抓住生命科技
跳动的脉搏
首款专注于tRNA研究的PCR芯片――nrStar™ Human tRNA Repertoire PCR Array
【字体: 大 中 小 】 时间:2016年12月08日 来源:康成生物
编辑推荐:
Arraystar最新发布了市场上首款专注于tRNA研究的PCR芯片――nrStar™ Human tRNA Repertoire PCR Array。该款芯片可同时检测66个细胞核tRNA和22个线粒体tRNA,覆盖了tRNA权威数据库GtRNAdb和tRNAdb中的所有反密码子,方便客户快速地进行tRNA表达谱分析。
转运RNA (tRNA) 是生物体内分布最广泛、含量最丰富的非编码小RNA分子。它携带并转运氨基酸,参与蛋白翻译,是连接mRNA与蛋白质的重要桥梁。细胞增殖[1]、分化[1, 2]和凋亡[3]等一系列生物学过程都伴随着tRNA水平的变化。反之,tRNA表达谱的改变也会影响细胞发育过程中的命运抉择。表达失调的tRNA可以促进肿瘤的发生和癌症进程[2, 4-11]。另外,许多其它疾病比如II型糖尿病[12]、亨廷顿症[13]以及HIV感染[14]都出现了tRNA表达与分布紊乱。tRNA研究已逐渐成为生物学过程和疾病研究的重要组成部分。
Arraystar最新发布了市场上首款专注于tRNA研究的PCR芯片――nrStar™ Human tRNA Repertoire PCR Array。该款芯片可同时检测66个细胞核tRNA和22个线粒体tRNA,覆盖了tRNA权威数据库GtRNAdb和tRNAdb中的所有反密码子,方便客户快速地进行tRNA表达谱分析,为下一步的密码子偏好性、蛋白翻译效率和准确性、细胞动力学以及病毒感染的细胞嗜性等研究提供重要线索,是进行tRNA研究必不可少的工具。
tRNA存在种类繁多的修饰,对其发挥功能十分关键。然而,这些修饰尤其是甲基化修饰会严重阻碍反转录的进行,导致cDNA合成终止或碱基错配。为此,Arraystar专门开发了针对tRNA的反转录试剂盒(rtStar™ tRNA-optimized First-Strand Synthesis Kit)。该试剂盒采用了一种高效的RNA去甲基化酶AlkB,能够有效的去除tRNA上的甲基化修饰,极大的提高cDNA合成质量。与此试剂盒组合使用,研究人员能够获得更为真实可靠的tRNA表达变化,为研究蛋白质组或tRNA来源片段(tRFs)提供重要信息。
为了保证数据可信度,芯片还包含了3个看家小RNA作为内参,以及3个质控对照RNA Spike-in、PCR阳性对照(PPC)和基因组DNA对照(GDC),分别用于监测cDNA合成质量、PCR效率和基因组DNA污染。
在后续研究中,敲低tRNA[15] 或过表达tRNA [2, 16, 17]可以进一步揭示特定tRNA在细胞生命活动中的作用。其它用于非编码RNA研究的相关技术也适用于tRNA的后续研究。
产品列表
|
产品名称 |
规格 |
描述 |
|
nrStar™ Human tRNA Repertoire PCR Array |
384-well (4*96) plate |
包括66个细胞核tRNA和22个线粒体tRNA |
芯片特点
• 囊括GtRNAdb和tRNAdb数据库中所有的细胞核与线粒体反密码子
• 伴随的去甲基化处理使得检测结果更加真实可靠
• 所有引物均在多种细胞和组织中通过验证
• 即拆即用型384孔板,几小时内便可得到结果
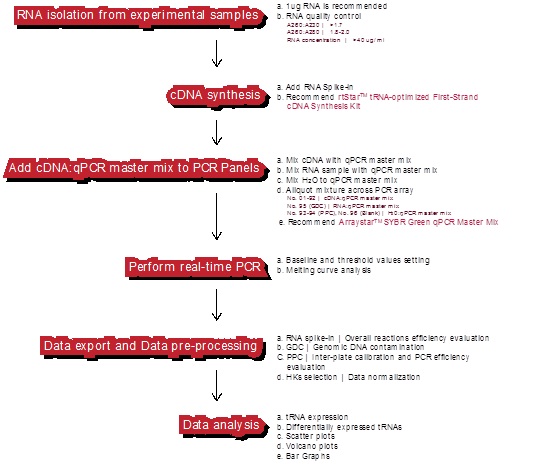
工作流程
适用实时定量荧光PCR仪:
ABI ViiA™ 7,ABI 7500 & ABI 7500 FAST,ABI 7900HT,ABI QuantStudio™ 6 Flex Real-Time PCR system,ABI QuantStudio™ 7 Flex Real-Time PCR system,ABI QuantStudio™ 12K Flex Real-Time PCR System,Bio-Rad CFX384,Bio-Rad iCycler & iQ Real-Time PCR Systems,Eppendorf Realplex,QIAGEN Rotor Gene Q100,Roche LightCycler 480,Stratagene Mx3000,Roche LightCycler 480
数据库
tRNA反密码子信息(GtRNAdb和tRNAdb数据库):
|
细胞核tRNA (66): Ala-AGC, Ala-CGC, Ala-GGC, Ala-TGC, Arg-ACG, Arg-CCG, Arg-CCT, Arg-TCG, Arg-TCT, Asn-ATT, Asn-GTT, Asp-ATC, Asp-GTC, Cys-GCA, Gln-CTG-1, Gln-CTG-2, Gln-TTG-1, Gln-TTG-2, Glu-CTC, Glu-TTC, Gly-CCC-1, Gly-CCC-2, Gly-GCC, Gly-TCC, His-GTG, Ile-AAT, Ile-TAT, Ini-CAT, Leu-AAG, Leu-CAA, Leu-CAG, Leu-TAA, Leu-TAG, Lys-CTT-1, Lys-CTT-2, Lys-TTT, Met-CAT, Phe-GAA, Pro-AGG, Pro-CGG, Pro-GGG, Pro-TGG, Sec-TCA, Ser-ACT, Ser-AGA, Ser-CGA, Ser-GCT, Ser-GGA, Ser-TGA, Sup-CTA, Sup-TTA, Thr-AGT-1, Thr-AGT-2, Thr-CGT, Thr-TGT-1, Thr-TGT-2, Trp-CCA, Tyr-ATA, Tyr-GAT, Tyr-GTA, Val-AAC, Val-CAC-1, Val-CAC-2, Val-CAC-3, Val-TAC-1, Val-TAC-2 |
|
线粒体tRNA (22): mt-Ala-TGC, mt-Arg-TCG, mt-Asn-GTT, mt-Asp-GTC, mt-Cys-GCA, mt-Gln-TTG, mt-Glu-TTC, mt-Gly-TCC, mt-His-GTG, mt-Ile-GAT, mt-Leu-TAA, mt-Leu-TAG, mt-Lys-TTT, mt-Met-CAT, mt-Phe-GAA, mt-Pro-TGG, mt-Ser-GCT, mt-Ser-TGA, mt-Thr-TGT, mt-Trp-TCA, mt-Tyr-GTA, mt-Val-TAC |
立即索取nrStar Human tRNA Repertoire PCR Array的详细资料
参考文献
[1] Gingold H, Tehler D, Christoffersen NR, Nielsen MM, Asmar F, Kooistra SM, et al. A dual program for translation regulation in cellular proliferation and differentiation. Cell 2014;158:1281-92.
[2] Pavon-Eternod M, Gomes S, Rosner MR, Pan T. Overexpression of initiator methionine tRNA leads to global reprogramming of tRNA expression and increased proliferation in human epithelial cells. Rna 2013;19:461-6.
[3] Mei Y, Stonestrom A, Hou YM, Yang X. Apoptotic regulation and tRNA. Protein & cell 2010;1:795-801.
[4] Berns A. A tRNA with oncogenic capacity. Cell 2008;133:29-30.
[5] Waldman YY, Tuller T, Sharan R, Ruppin E. TP53 cancerous mutations exhibit selection for translation efficiency. Cancer research 2009;69:8807-13.
[6] Kushner JP, Boll D, Quagliana J, Dickman S. Elevated methionine-tRNA synthetase activity in human colon cancer. Proceedings of the Society for Experimental Biology and Medicine Society for Experimental Biology and Medicine 1976;153:273-6.
[7] Marshall L, Kenneth NS, White RJ. Elevated tRNA(iMet) synthesis can drive cell proliferation and oncogenic transformation. Cell 2008;133:78-89.
[8] Pavon-Eternod M, Gomes S, Geslain R, Dai Q, Rosner MR, Pan T. tRNA over-expression in breast cancer and functional consequences. Nucleic acids research 2009;37:7268-80.
[9] Zhou Y, Goodenbour JM, Godley LA, Wickrema A, Pan T. High levels of tRNA abundance and alteration of tRNA charging by bortezomib in multiple myeloma. Biochemical and biophysical research communications 2009;385:160-4.
[10] Begley U, Sosa MS, Avivar-Valderas A, Patil A, Endres L, Estrada Y, et al. A human tRNA methyltransferase 9-like protein prevents tumour growth by regulating LIN9 and HIF1-alpha. EMBO molecular medicine 2013;5:366-83.
[11] Goodarzi H, Nguyen HC, Zhang S, Dill BD, Molina H, Tavazoie SF. Modulated Expression of Specific tRNAs Drives Gene Expression and Cancer Progression. Cell 2016;165:1416-27.
[12] Krokowski D, Han J, Saikia M, Majumder M, Yuan CL, Guan BJ, et al. A self-defeating anabolic program leads to beta-cell apoptosis in endoplasmic reticulum stress-induced diabetes via regulation of amino acid flux. The Journal of biological chemistry 2013;288:17202-13.
[13] Girstmair H, Saffert P, Rode S, Czech A, Holland G, Bannert N, et al. Depletion of cognate charged transfer RNA causes translational frameshifting within the expanded CAG stretch in huntingtin. Cell reports 2013;3:148-59.
[14] van Weringh A, Ragonnet-Cronin M, Pranckeviciene E, Pavon-Eternod M, Kleiman L, Xia X. HIV-1 modulates the tRNA pool to improve translation efficiency. Molecular biology and evolution 2011;28:1827-34.
[15] Fu G, Xu T, Shi Y, Wei N, Yang XL. tRNA-controlled nuclear import of a human tRNA synthetase. The Journal of biological chemistry 2012;287:9330-4.
[16] Gong M, Gong F, Yanofsky C. Overexpression of tnaC of Escherichia coli inhibits growth by depleting tRNA2Pro availability. Journal of bacteriology 2006;188:1892-8.
[17] Yona AH, Bloom-Ackermann Z, Frumkin I, Hanson-Smith V, Charpak-Amikam Y, Feng Q, et al. tRNA genes rapidly change in evolution to meet novel translational demands. eLife 2013;2:e01339.




